REVIEW ARTICLE
SANTOS, Patrícia Marques [1], NASCIMENTO, Marcelo Trindade [2]
SANTOS, Patrícia Marques. NASCIMENTO, Marcelo Trindade. Biotic homogenization in tree communities of tropical forests: a systematic review. Revista Científica Multidisciplinar Núcleo do Conhecimento. Year: 08, Issue: 08, Volume: 04, pp. 50-77. August 2023. ISSN: 2448-0959, Access link: https://www.nucleodoconhecimento.com.br/biology/biotic-homogenization, DOI: 10.32749/nucleodoconhecimento.com.br/biology/biotic-homogenization
ABSTRACT
A systematic literature review was conducted on biotic homogenization in tropical forests and its consequences for the diversity of tree communities. We evaluated articles indexed in Scopus® and Web of Science® from 1945 to 2022. The selection of publications was carried out using the Rayyan platform. The bibliometric method used for database mapping was co-word analysis in VOSviewer software. We distinguished three lines of research on biotic homogenization: invasion ecology of species; environmental and anthropogenic factors; and functional and phylogenetic diversity. The most studied and perhaps the most affected component was the taxonomic component. Biotic homogenization events are widely distributed throughout the tropical region, with the most affected forest type being tropical rainforest, and the main cause identified was land use change related to anthropogenic degradation. Studies on biotic homogenization in the tropics have intensified in the last decade and have resulted in significant advances in understanding this process, such as the realization that hyperdiverse environments are susceptible to biotic homogenization, which can be mediated by generalist native species. We observed the emergence of another component of diversity to be included in the homogenization concept: homogenization of ecological interactions.
Keywords: Bibliometrics, Conservation, Fragmentation, Tropical forests, Biotic homogenization.
1. INTRODUCTION
The increase in similarity among biological communities is the main characteristic of biotic homogenization (biological homogenization). This process, first described by Elton (1958), expressed concern about the spread of exotic species. The term “biotic homogenization” was defined later by Mckinney and Lockwood (1999), where the authors highlighted two fundamental factors for this process: land use change and the introduction of exotic species. These authors, when proposing this definition, mainly addressed taxonomic homogenization, a concept that has been evolving since then. Rahel (2000) added the time factor to the concept, defining it as the change in similarity between communities over time. Olden and Poff (2004) included the multidimensional character in the assessment of the process, incorporating functional and genetic components. Currently, biotic homogenization is defined as the alteration of previously distinct biotas that have become more similar over time at different levels of organization, e.g., genetic, taxonomic, and functional (OLDEN; ROONEY, 2006).
The intensification of human activities in general and, in some cases, reaching remote and previously undisturbed areas, has facilitated the invasion of exotic species into previously inaccessible areas, breaking natural biogeographic barriers and thus favoring biotic homogenization (CHARLES S. ELTON, 1958; KOLAR; LODGE, 2001; KRAMER; ZWIENER; MÜLLER, 2022).
All major causes of biodiversity loss on the planet are in some way linked to human actions (e.g., deforestation, habitat loss, fragmentation, pollution of water bodies, hunting, introduction of exotic species, climate change) and their secondary effects (DÍAZ et al., 2019; ESTES et al., 2011; FINLAYSON et al., 2005; LEWIS; EDWARDS; GALBRAITH, 2015; LOBO et al., 2011; MAXWELL et al., 2016; PENN; DEUTSCH, 2022). The interaction of different human actions can intensify their effects on biodiversity loss, leading to biotic homogenization in previously highly diverse areas, such as biodiversity hotspots worldwide (SILVA; TABARELLI, 2000; ZWIENER, 2018), and tropical forests (DIRZO, 2001; PAIN et al., 2021).
The exponential increase in the number of publications has brought great challenges for review studies. The application of systematic review and bibliometric methods allows dealing with this wealth of data, filtering important works by estimating their impact and discovering underlying patterns in a field of knowledge (ZUPIC; ČATER, 2015). Thus, the objective of this study was to conduct a systematic literature review on biotic homogenization and its consequences on the diversity of tropical tree communities, highlighting the main knowledge gaps.
2. MATERIALS AND METHODS
2.1 SYSTEMATIC LITERATURE REVIEW
We conducted a systematic literature review based on Moher et al. (2009). We evaluated scientific articles that address aspects related to biotic homogenization in tropical forests. The search for publications was performed without temporal limitations and language restrictions to obtain all available publications in the databases (1945 to 2022). Based on this premise, a bibliographic search was carried out in two of the main academic databases: Scopus® and Web of Science®.
To search for articles, we considered the following terms related to the topic: “homogenization” OR “tree homogenization” OR “plant homogenization” OR “biotic homogenization” AND “plant”; which should be present in the title, abstract, or keywords of the articles. The terms used in this search were selected from reference articles in the field, and the searches were conducted up to June 2022.
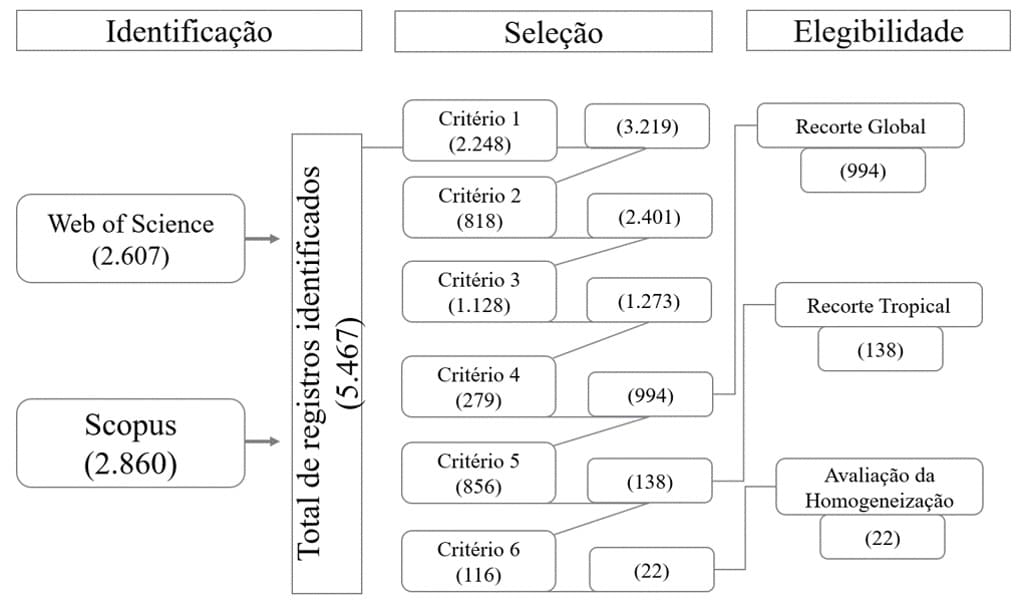
We selected documents using screening criteria adapted from Moher et al. (2009) and O’Dea et al. (2021), as shown in Figure 1. As the first criterion, we selected only documents from knowledge areas related to environmental and earth sciences. In the second criterion, we excluded duplicate articles from the databases, considering only one document. As the third criterion, we considered only articles that dealt with biotic homogenization as a central theme. The fourth criterion considered only articles that studied tree species. In the fifth criterion, we selected only articles from tropical regions, and in the sixth and final criterion, we selected only articles that tested the process of biotic homogenization as described by Olden and Rooney (2006). These authors identified four routinely used ways to test biotic homogenization: 1- “Species pool” studied at two time periods; 2- Records of existing and reconstructed historical species occurrences; 3- “Species pool” of existing and reconstructed historical species (corrected for extinctions); 4- Existing species sets – community similarity. Articles that only mentioned the process without conducting an experiment or evaluation to prove the phenomenon were not considered. The entire process of filtering publications was carried out on the Rayyan selection platform (OUZZANI et al., 2016).
Figure 1. Flowchart describing the search protocol for documents selected in the systematic review from 1945 to 2022. 1st Criterion – Knowledge areas; 2nd Criterion – Duplicates; 3rd Criterion – Biodiversity loss; 4th Criterion – Plants as the central theme; 5th Criterion – Only articles from tropical regions; 6th Criterion – Only articles that quantified biological homogenization
2.2 DATABASE ANALYSIS
From the articles selected for the final analysis, we extracted the following data: year, authors, assessment scale (local, regional, global), field of knowledge, studied biome, vegetation type, study category (observational, experimental, others), biodiversity component analyzed (Taxonomic, Functional, and Phylogenetic), approach and metric used, evaluation period, and main cause identified for biotic homogenization.
2.3 BIBLIOMETRIC METHODS
The bibliometric method used to map the database was co-word analysis (VAN ECK; WALTMAN, 2014). This analysis evaluates the full text of publications and creates maps based on distance.
Each term is sized based on its relevance in a particular dataset, and the thickness of the lines connecting them is determined by the strength of association (frequency at which the terms were found together). The distance between two items reflects the strength of the relationship between them. We selected terms with a minimum of two occurrences and then narrowed down to the top 100 terms, following Van Eck and Waltman (2010). The software used to map co-words was VOSviewer (JAN; LUDO, 2010). The workflow used in this study was described by Zupic and Carter (2015).
3. RESULTS
A total of 5,467 articles published from 1945 to 2022 were identified. After the filtering process, 994 articles were considered for the analysis of biotic homogenization on a global scale, 138 on a tropical scale, and of these, only 22 tested biotic homogenization (Figure 1).
3.1 CHARACTERIZATION OF BIOTIC HOMOGENIZATION PUBLICATIONS IN THE TROPICS
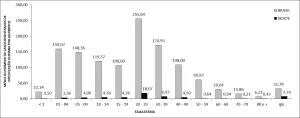
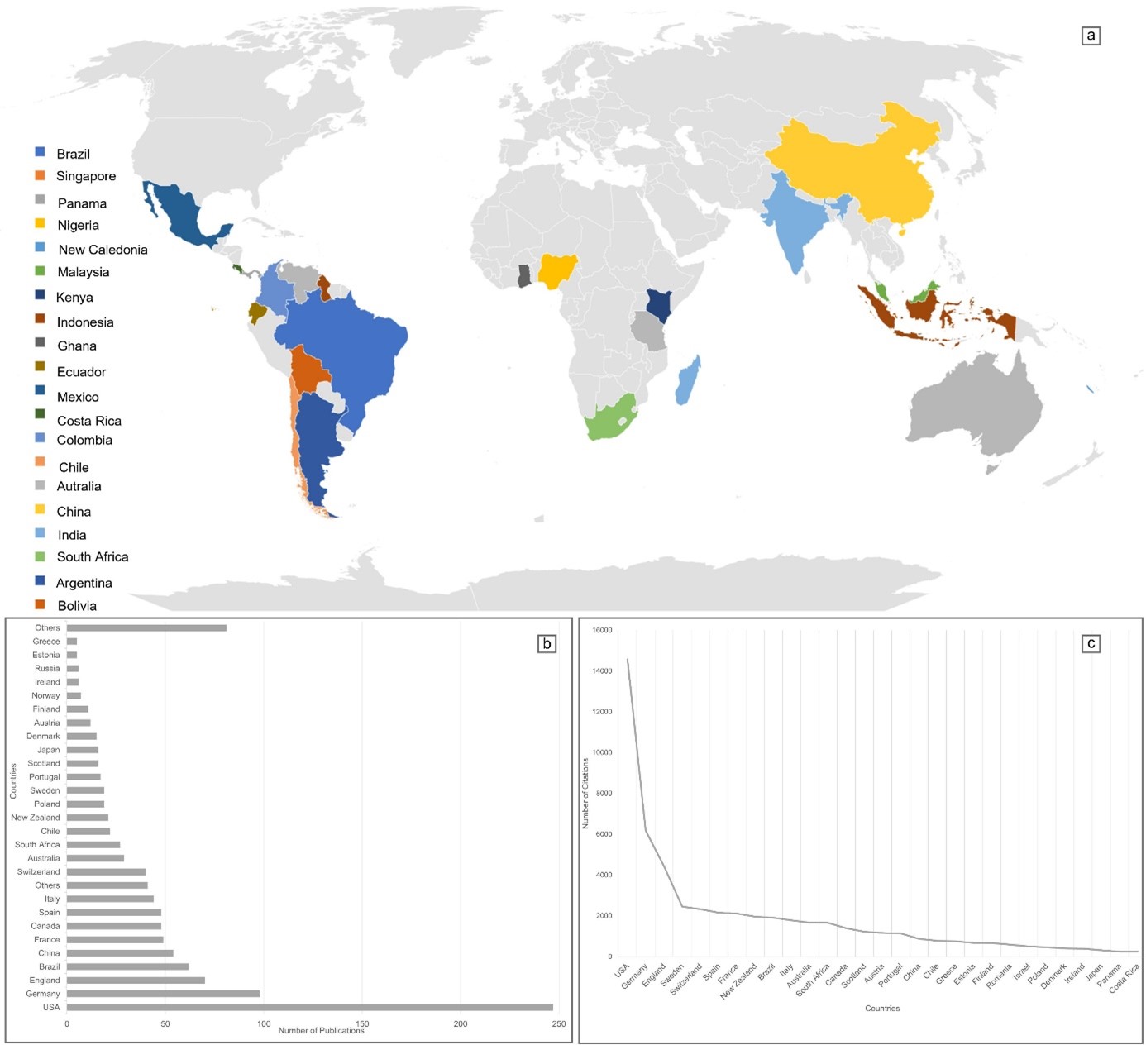
In the tropical scale cut-off (n = 138) shown in Figure 1, we observed that twenty-six countries (Figure 2) have records of biotic homogenization, with China (n = 54), Brazil (n = 31), and Australia (n = 29) standing out (Figure 2b). In the tropical region, the countries with the most citations in these publications are Brazil (n = 1,400), followed by Mexico (n = 313) and Costa Rica (n = 247) (Figure 2c).
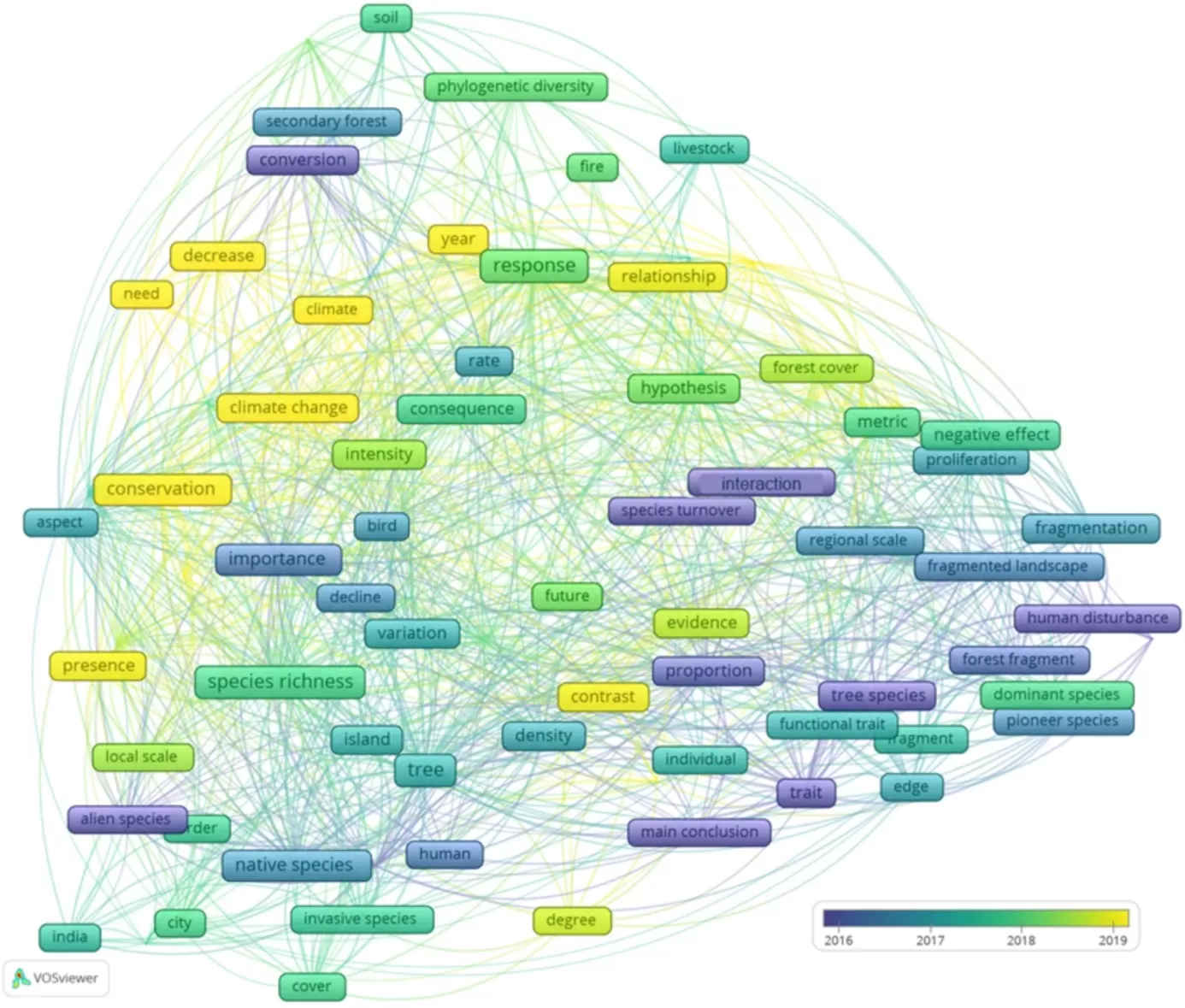
Figure 2. Temporal evaluation of co-words extracted from the text of titles, author-assigned keywords, and abstracts of articles selected in the systematic review on biotic homogenization from 1945 to 2022. Evaluation based on 994 articles on the selected theme. The color scale represents the time variation, with yellow terms originating from more recent publications and blue terms from older ones
The co-word analysis for the tropics highlighted three research lines based on biotic homogenization: ecology of species invasion (green); environmental and anthropogenic factors, and functional diversity (red); and phylogenetics (blue), as shown in Figure 3. Furthermore, the underlying terms emphasize the importance of regional themes such as landscape context and species interactions with the environment.
Figure 3. Co-word analysis extracted from the text of titles, author-assigned keywords, and abstracts of articles filtered for the tropics in the systematic review on biotic homogenization from 1945 to 2022. Evaluation based on 138 articles on the selected theme narrowed down to the tropics. The colors represent the clustering of research lines: Phylogenetic diversity (blue); ecology of species invasion (green); environmental and anthropogenic factors and functional diversity (red)
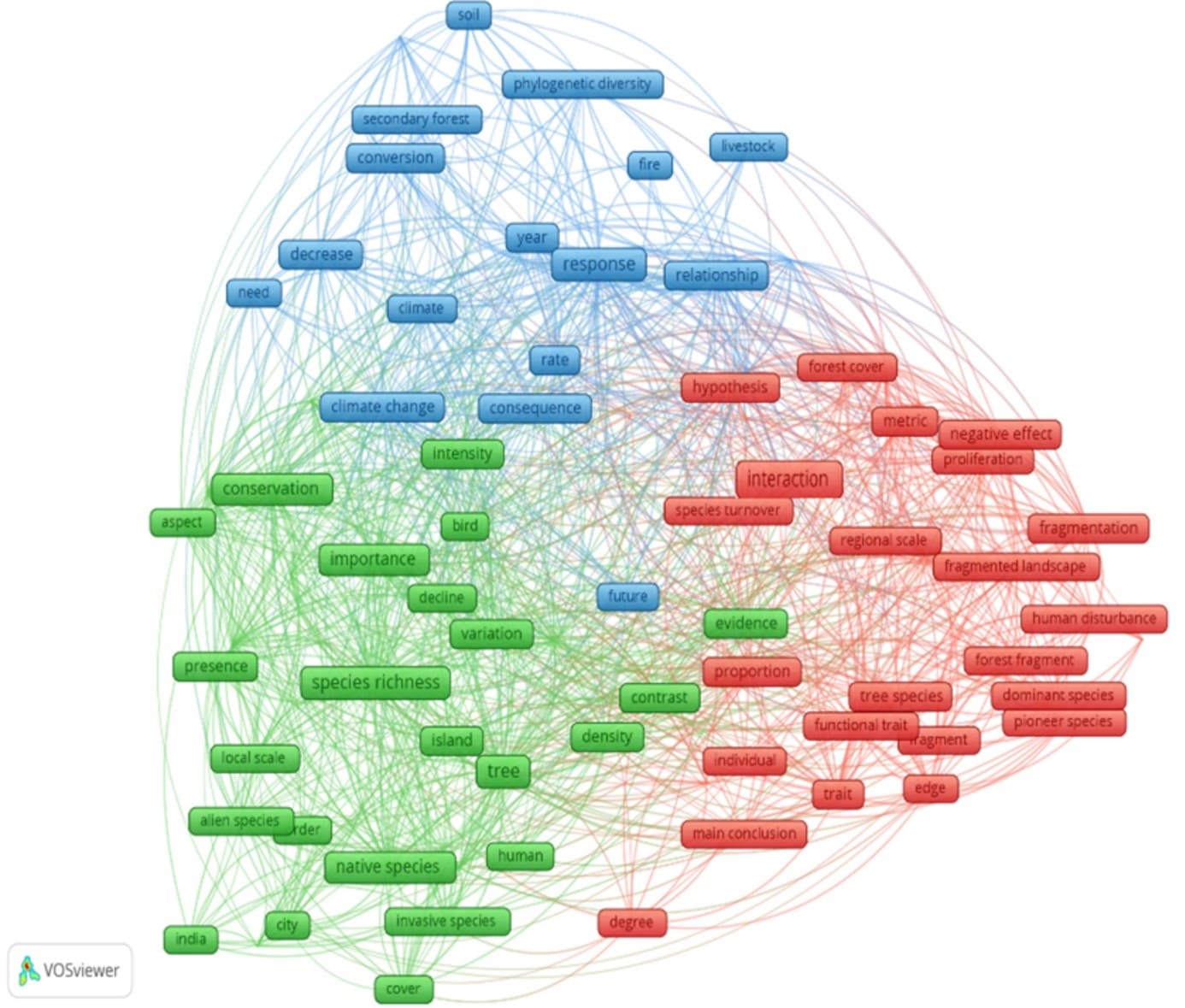
In the red group, we observe terms such as interaction, species turnover, and species substitution related to fragmentation, highlighting a new trend also noted in the deeper analysis of these publications, which is the study of the homogenization of interactions. These analyses have been more recently addressed due to a growing concern about the loss of key species observed in the areas, which has promoted homogenization in interactions.
The study of the dynamics of research lines over time showed that concepts like phylogenetic diversity and climate change are being applied more recently (Figure 4), while the interaction of anthropogenic actions (fragmentation, habitat loss, urbanization) with species composition are themes with a longer history of study, as well as invasion ecology (introduction of exotic species, characteristics of these species, and the environment).
Figure 4. Temporal evaluation of co-words extracted from the text of titles, author-assigned keywords, and abstracts of articles filtered for the tropics in the systematic review on biotic homogenization from 1945 to 2022. Evaluation based on 138 articles on the selected theme. The color scale represents the temporal variation, with yellow terms originating from more recent publications, while blue terms are from older ones
3.2 CHARACTERIZATION OF PUBLICATIONS THAT TESTED BIOTIC HOMOGENIZATION
Publications that assessed biotic homogenization in the tropics (n = 22) revealed that 64% utilized spatial scale, while only 36% tested through a temporal assessment. Despite the small number of publications that effectively tested homogenization, studies were registered in nine countries in the tropical region, with Brazil (n = 9), India (n = 3), and Mexico (n = 2) standing out, as seen in Figure 5a and b.
Figure 5. Geographic distribution of publications by countries in the tropical region (a) and number of publications (b) obtained in the systematic review on Biotic Homogenization from 1945 to 2020
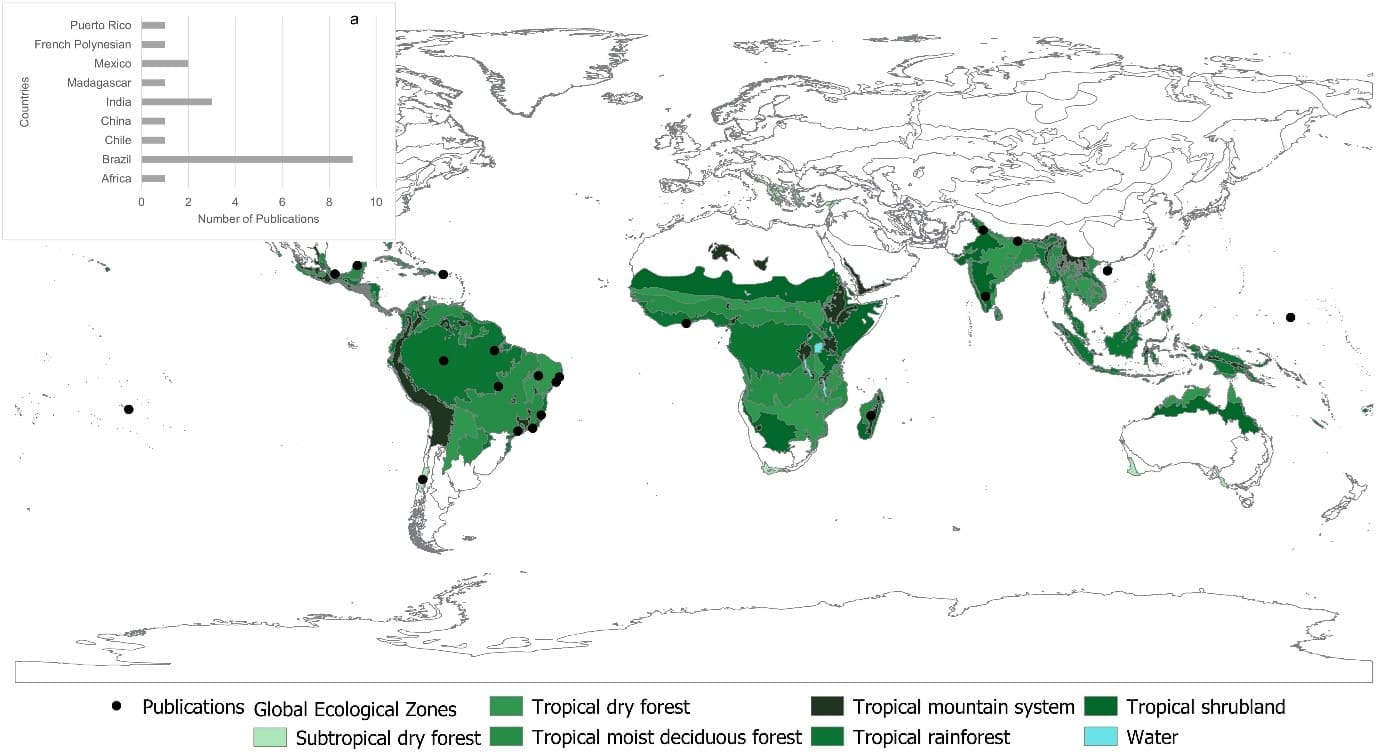
The tropical rainforest type (dense ombrophilous forest) gathers the highest number of studies in the tropics (n = 14), followed by the tropical semideciduous forest (n = 2), with the remaining occurrences classified in other ecological zones (Figure 5a). In the case of Brazil, the biome with the highest number of publications on the subject was the Atlantic Forest (n = 5), followed by the Amazon Rainforest (n = 2), with the Caatinga and the Amazon/Cerrado transition area each presenting one publication (Figure 5a).
Among the methods identified by Olden and Rooney (2006), the most used by the authors was the comparison between communities (n = 14), followed by temporal scale assessment (Figure 6). Despite the higher number of publications using the comparison between communities, proportionally, the evaluation at a temporal scale was more successful, confirming 75% (n = 6), compared to 64% (n = 9) for the method using the comparison between communities.
Figure 6. Flowchart describing the methods, assessment, and main causes identified for the occurrence of Biotic Homogenization (HB) in publications that tested this phenomenon (n=21). The asterisk indicates the main causes for the differentiation of communities.
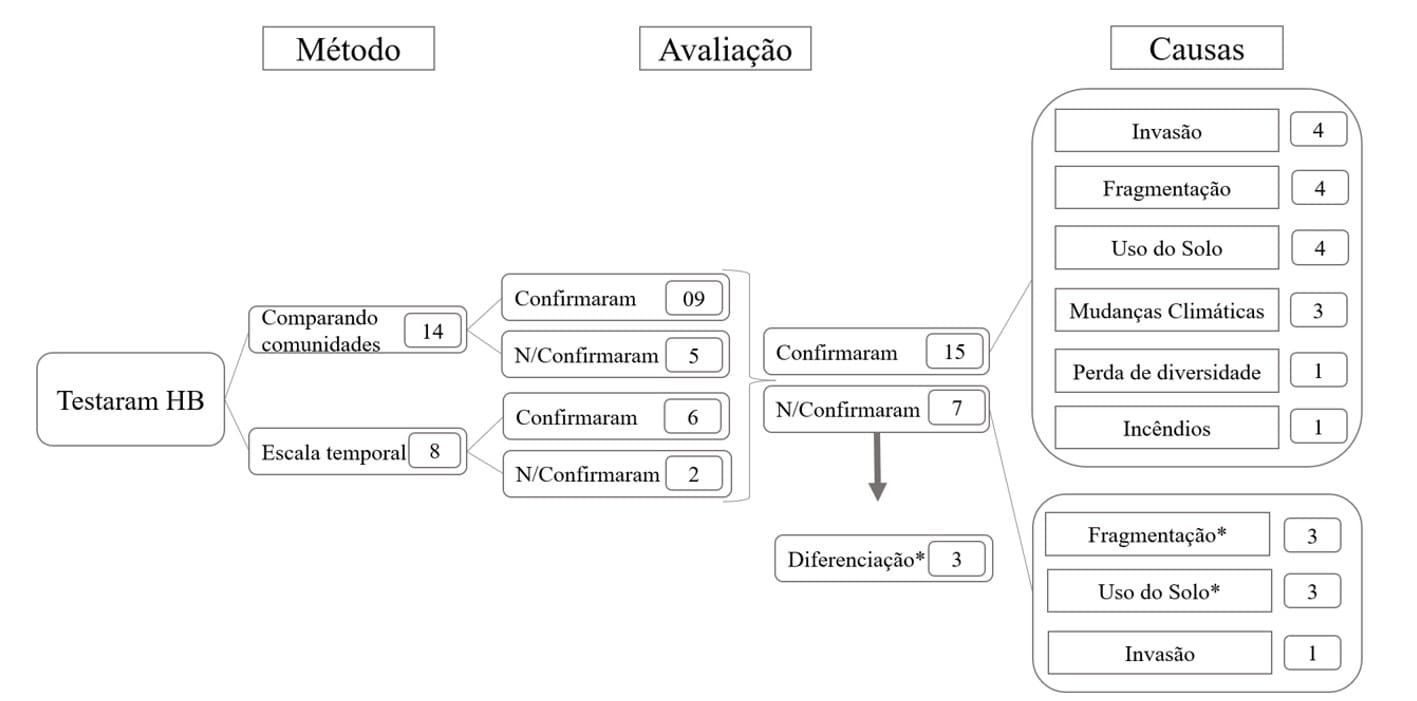
Evaluating the publications that tested the occurrence of this phenomenon (n = 22), it became evident that the main tested causes for biotic homogenization were Fragmentation (n = 4), Species Invasion (n = 4), and Land Use Change (n = 4).
However, Arroyo-Rodríguez et al. (2013), Solar et al. (2015), Sfair et al. (2016), Inague; Zwiener; Marques (2021) observed the opposite phenomenon, i.e., an increase in biotic diversity in the areas. This process seems to be scale-dependent, as they observed differentiation at the local level (plot, fragment) while at the landscape scale, this difference was not confirmed. The diversification in these areas resulted from fragmentation (MARTINEZ et al., 2010); multiple human disturbances (SFAIR et al., 2016); land use history (URIBE et al., 2021); and climate change (INAGUE; ZWIENER; MARQUES, 2021).
Among the publications that analyzed the variation in community similarity on a temporal scale (n = 8), i.e., before and after the occurrence of the degradation event, as suggested by Olden and Rooney (2006), two did not confirm biotic homogenization. Lobo et al. (2011), in a comparative analysis of the tree flora of the Atlantic Forest before and after 1980, observed that the plant community underwent a process of fragmentation and a consequent increase in the number of native pioneer and early secondary species.
The increase in floristic similarity as a result of environmental degradation events (forest fires) was also reported by Silva et al. (2000). Over a seven-year temporal scale (2007 to 2014), it was observed that the increase in fire frequency led to the local extinction of species and, consequently, an accelerated increase in similarity, in a particularly sensitive area such as the transition between the Cerrado and the Amazon Forest.
Homogenization was also assessed as a result of climate change (INAGUE; ZWIENER; MARQUES, 2021; ZWIENER, 2018). In this context, current climatic conditions were compared with future scenarios. These authors demonstrated that under severe climate change scenarios, there would be a large-scale reduction in diversity and an increase in species similarity among communities. They concluded that the expansion of generalist species would be responsible for the increase in species similarity. Evaluating the coastal vegetation of the Atlantic Forest, Inague; Zwiener; Marques (2021) found diversification in the taxonomic component and homogenization in the functional component, highlighting that climate change would restrict functional diversity to more acclimatized traits.
Another factor addressed on a temporal scale was the consequences of extreme drought events on taxonomic, functional, and phylogenetic diversity in humid forests (AGUIRRE-GUTIÉRREZ et al., 2020). The temporal scale used covers the period from 1980-1990 to 2010-2013, observing an increase in taxonomic and functional diversity in humid forests, while drier forests showed a reduction. In phylogenetic diversity, a significant reduction was observed from wetter to drier gradients.
A recurring factor in evaluating homogenization is land use and land cover change; however, only one publication assessed it on a temporal scale. Uribe et al. (2021) studied the influence of this factor on biodiversity in Pinus plantations, with the temporal scale represented by the rotation time of each planting (20 years). It was found that plantations that directly replaced native forests had a large number of specialist species and a lower number of generalists, and the number of rotations did not influence local diversity. Thus, land use and land cover change did not promote species homogenization in the Pinus plantation area.
Publications that tested the occurrence of the homogenization process and found negative results evaluated the causes: fragmentation, land use change, and biological invasion. Among them, Martines et al. (2010) evaluated land use change, environmental characteristics, and the presence of invasive species, concluding that species establishment was related to the substrate and not to land use change. Sfair et al. (2016) evaluated whether homogenization or taxonomic/functional divergence was occurring in edge-effect areas and homogenization was not confirmed. Arroyo-Rodrigues et al. (2013) also evaluated these two processes in a deforestation gradient and did not observe homogenization.
Meyer et al. (2014) investigated the propensity of island environments for homogenization by evaluating land use change and species invasion. It was found that these factors caused significant changes in the environments but did not result in homogenization of the studied areas.
Sanaphre-Villanueva et al. (2017) evaluated whether fragmentation would promote homogenization in taxonomic and functional diversity of generalist species in a successional chronosequence. These authors observed taxonomic homogenization as a result of the dominance of generalist species but concluded that this dominance did not result in functional homogenization.
The most addressed component of biodiversity in the research was the taxonomic component (n = 13). Some studies addressed two components simultaneously (taxonomic and functional (n = 5)), and only two articles evaluated all three components (taxonomic, functional, and phylogenetic). Multi-taxa evaluation was only performed in three publications (ARAVIND et al., 2010; SOLAR et al., 2015; ZHANG et al., 2022); of these, two confirmed the occurrence of homogenization, and one identified diversification.
Table 1. List of publications that tested biotic homogenization in the tropical region, with a focus on the main characteristics addressed. In Gray are the publications that did not confirm biotic homogenization. Functional characteristics evaluated: total leaf area (LA), minimum photosynthetic unit (MPU), leaf dry matter content (LDMC), specific leaf area (SLA); wood specific gravity (WSG); leaf pulvinus (LPulv); leaf pubescence (LPb); plant exudates (Ex); plant spines (Sp); leaf composition (LC); dispersal syndrome (Dis); deciduousness (LD); seed volume (SV); Maximum Height (Hmax); Diameter at Breast Height (DAP); adult body mass (MCA); diet breadth (AD); habitat breadth (AH); trophic level (NT); activity cycle (Cat); foraging stratum (EF); migratory status (SM); lifespan (TV); growth form/life form (FC); and shoot length (CB); stem specific conductivity potential (kp), vessel lumen fraction (VLF), vessel diameter (VD), vessel density (pV), leaf nitrogen content (NL), and leaf phosphorus content (PL), leaf thickness (ThicknessL), maximum rates of carbon assimilation in light-saturated carbon assimilation rates (Asat); wood density (WSG); phenology (F); guild (G) and nitrogen-fixing capacity (CFN)
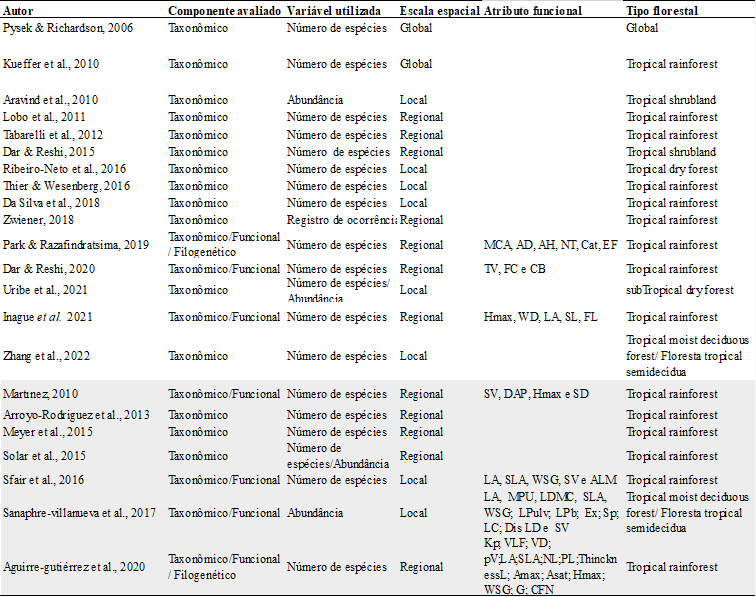
Among the set of articles that used only the taxonomic component to assess biotic homogenization (n = 15), a large portion concluded that homogenization is occurring (n=10); two publications did not confirm biotic homogenization (SOLAR et al., 2015; URIBE et al., 2021). Four publications were inconclusive in their results (ARROYO-RODRÍGUEZ et al., 2013; MEYER et al., 2014; PYSEK; RICHARDSON, 2006; URIBE et al., 2021).
The functional component is being evaluated in biotic homogenization research through the following traits: dispersal syndromes (n = 2), life forms (n = 2), leaf characteristics (n = 7), and wood density (n = 3). The chosen traits are related to water and nutrient economy, shade tolerance, and colonization. That is, to the ability to survive in a restrictive environment, as would be expected in degraded areas or under biological invasion.
Among the publications that assessed the functional component (n = 7), four of these articles found evidence of functional homogenization. The rest identified functional diversification, and the justification in these works was the great functional redundancy of the areas, which until then maintained the functional diversity of the region.
Only two studies addressed phylogenetic homogenization (AGUIRRE-GUTIÉRREZ et al., 2020; PARK; RAZAFINDRATSIMA, 2019) by jointly assessing taxonomic and functional diversity. These works showed the importance of simultaneously approaching the three components due to the uniqueness of each component and its differentiated effects. These studies concluded that biotic homogenization is occurring in the evaluated areas and found homogenization in all three components of biodiversity. Only one of these articles evaluated on a temporal scale.
4. DISCUSSION
One of the first observations from this review is the generality in the use of the term ‘biotic homogenization,’ often used merely to denote the loss of diversity in a region, without confirming the process. This simplistic use of the term was present in several articles dealing with species richness and diversity or those addressing the diversity of an area, noting the invasion of a species and warning of the possibility of future biotic homogenization without actually testing homogenization, as emphasized by Olden and Rooney (2006).
The inappropriate use may result from the late definition of the biotic homogenization process, which was officially defined only in 1999 (MCKINNEY; LOCKWOOD, 1999). Although documented since 1958 when Charles S. Elton described the biotic homogenization process resulting from the invasion by exotic species. Or even the generality of the term used to define the process, as discussed by Olden and Rooney (2006).
Since its official definition by Mckinney and Lockwood (1999), successive records have highlighted the biotic homogenization process as an unavoidable effect of biological invasion (LAMBDON et al., 2008). However, as highlighted by Oldem (2006), invasion is not biotic homogenization. Not all invasions lead to biotic homogenization or species extinction, and not all variation in species diversity corresponds to biotic homogenization (ARROYO-RODRÍGUEZ et al., 2013).
The development of the research area, as observed, reflects exactly the history of biotic homogenization research, which initially focused on homogenization, strongly related to the expansion of exotic species and their characteristics (CHARLES, 1958; MCKINNEY, 1998), and later incorporating other facets of diversity (GARCÍA-NAVAS et al., 2020; OLDEN; POFF, 2004), broadening the evaluation scale both in space and time (RAHEL, 2002), and more recently relating to current issues such as habitat fragmentation and climate change (AGUIRRE-GUTIÉRREZ et al., 2020; INAGUE; ZWIENER; MARQUES, 2021). The low number of articles that tested biotic homogenization in the tropical region (n = 22) shows the intrinsic complexity of this type of analysis. Since it is essentially a comparative analysis, it must be done on a temporal scale, based on consistent data from two or more different areas, with at least one area free from impacts.
The difference in the number of biotic homogenization records in temperate regions (n = 994) versus the tropics (n = 138) can be justified by three issues: the early onset of environmental degradation during the industrial revolution (FOSTER, 1999), sampling bias, and the number of publications found for the temperate region, which is more than triple the number of articles recorded for the tropics (n = 856). Another factor is the lack of standardized long-term field data to effectively test biotic homogenization in the tropical region (CONDIT, 1995; HARRIS et al., 2021).
Environmental degradation was a prominent factor in homogenization studies, being the main cause pointed out for its occurrence in the evaluated publications, varying only in the scale of local or regional degradation (e.g., land use change, fragmentation, fire) and global (e.g., climate change).
In Brazil, studies mainly address the effects of human action, historically present in the Atlantic Forest and in the Amazon/Cerrado transition, a region now known as the deforestation arc, the last agricultural frontier in central Brazil (FEARNSIDE, 2020).
The prevalence of the Atlantic Forest Biome in studies that tested biotic homogenization in the tropics (25% of the scientific production in the area) seems to be related to intrinsic factors such as historical fragmentation in this biome (MELLO et al., 2020), and the high biodiversity and endemism (MITTERMEIER et al., 2004). In addition, this region has the highest concentration of research institutions and human resources active in Brazil (INEP, 2022).
We observed the dominance of studies based only on the taxonomic component, probably due to the cost involved in research with functional and phylogenetic data collection and the limited availability of robust databases for these components in the tropics (GUIMARÃES et al., 2005).
The high richness of the tropical region allows for the coexistence of many animal and plant species and an immeasurable number of interactions, such as pollination, herbivory, dispersal, and nutrient transfer (TOBY KIERS et al., 2010). Thus, these interactions are threatened by human actions and the increase in biotic homogenization, which has led to the breakdown of these plant-animal relationships, promoting the homogenization of interactions (CÂMARA et al., 2018; OLIVEIRA et al., 2019; TYLIANAKIS et al., 2007). This process was detected in the assessment of the database of this article and can be considered one of the current trends in the study of homogenization.
As important as evaluating the various facets of biodiversity of a specific plant community or region is to evaluate the largest number of taxa simultaneously, since each one can respond differently to the biotic homogenization process due to the cascading effect of diversity loss in one of these taxa, as observed in the studies of (ARAVIND et al., 2010; PARK; RAZAFINDRATSIMA, 2019; SOLAR et al., 2015; ZHANG et al., 2022).
Vegetation communities are highly dynamic, mainly under the influence of human action (LAURANCE et al., 2006), making it almost impossible to predict the paths of the species replacement process. The studies bring two possibilities: the occurrence of biotic homogenization or diversification (ESCOBAR et al., 2013). And, as discussed earlier, long-term studies are fundamental to assess changes in communities over time and thus improve our understanding of homogenization versus differentiation processes in landscapes modified by humans (HADDAD et al., 2015; KRAMER et al., 2022; REES et al., 2001).
Based on the studies that tested biotic homogenization in the tropics, we realize that the loss of species diversity is occurring in many cases, but not always resulting in biotic homogenization. However, non-confirmation may also result from the approach used to assess homogenization. Most of the studies highlighted here used similarity between communities or disturbance gradients, as described by Olden and Rooney (2006). However, as discussed by these authors, this is not the best way to quantify biotic homogenization; it is preferable to evaluate at two points in time (OLDEN; ROONEY, 2006).
5. FINAL CONSIDERATIONS
Studies on biotic homogenization in the tropics have provided significant advancements in understanding this process. The successive yet persistent and large-scale degradation can lead highly diverse regions such as hotspots to biotic homogenization.
We found that unlike the initial concept that placed the introduction of exotic species as the main cause of homogenization, more recently, publications have shown that the primary cause has been land use change, and in the tropics, native generalist species can play a significant role in homogenization through a significant increase in the dominance of these species in degraded environments.
Among the components commonly evaluated, the taxonomic component was the most affected, followed by the functional and phylogenetic components. However, these assessments were mainly conducted at a spatial scale. Few studies used a temporal scale to evaluate this phenomenon.
In addition to the taxonomic, functional, and phylogenetic components, the effect on ecological interactions or the homogenization of interactions is another facet of diversity to be evaluated, proving to be an emerging theme in the realm of biotic homogenization studies. The loss of these interactions represents a significant risk to the maintenance of ecosystem services.
For the success of conservation strategies in the future, it is important to understand the factors and patterns of biodiversity loss at spatial and temporal scales, and understanding biotic homogenization becomes essential in this context. This is a process that encompasses many dimensions of the modern biodiversity crisis, as it involves species invasions and extinctions.
Research on biotic homogenization should advance by increasing studies in priority conservation areas (biodiversity hotspots in the tropics). In addition, long-term monitoring studies should be implemented in areas where there are reports of species invasions to provide an appropriate assessment of the occurrence or absence of biotic homogenization.
Research on biotic homogenization is still very recent and therefore raises more questions than answers. It is necessary to clarify, for example, whether the tropical region is more susceptible to biotic homogenization and whether this process is faster or slower than observed in temperate areas.
Understanding the current scenario and making projections about the effects of habitat destruction and climate change on the process of biotic homogenization in priority conservation areas is fundamental for establishing mitigating actions.
ACKNOWLEDGMENTS
This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Funding Code 001, the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and the Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (FAPES). Marcelo Trindade Nascimento is supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico do Brasil (CNPq: 305617/2018-4) and the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ E-26/202.855/2018).
We thank Cristiane Marques Santos and John DuVall Hay for their critiques and suggestions on the manuscript.
REFERENCES
AGUIRRE-GUTIÉRREZ, J. et al. Long-term droughts may drive drier tropical forests towards increased functional, taxonomic and phylogenetic homogeneity. NATURE COMMUNICATIONS, p. 1–10, 2020.
ARAVIND, N. A. et al. Impact of the invasive plant, Lantana camara, on bird assemblages at Male Mahadeshwara Reserve Forest, South India. TROPICAL ECOLOGY, v. 51, n. 2, SI, p. 325–338, 2010.
ARROYO-RODRÍGUEZ, V. et al. Plant b-diversity in fragmented rain forests: testing floristic homogenization and differentiation hypotheses. Jornal of Ecology, v. 101, n. 6, p. 1449–1458, 2013.
CÂMARA, T. et al. Effects of chronic anthropogenic disturbance and rainfall on the specialization of ant–plant mutualistic networks in the Caatinga, a Brazilian dry forest. Journal of Animal Ecology, v. 87, n. 4, p. 1022–1033, 2018.
CHARLES S. E. The Ecology of Invasions by Animals and Plants. 1. ed. Chicago: Methuen & Co. Ltd. v. 1, 1958.
CONDIT, R. Research in large, long-term tropical forest plots. Trends in Ecology & Evolution, v. 10, n. 1, p. 18–22, 1995.
DÍAZ, S. et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science, v. 366, n. 6471, 2019.
DIRZO, R. Tropical Forests BT – Global Biodiversity in a Changing Environment: Scenarios for the 21st Century. In: CHAPIN, F. S.; SALA, O. E.; HUBER-SANNWALD, E. (Eds.). New York, NY: Springer New York, p. 251–276, 2001
ESCOBAR, F. et al. Plant b-diversity in fragmented rain forests: testing floristic homogenization and differentiation hypotheses. Jornal of Ecology, v. 101, p. 1449–1458, 2013.
ESTES, J. A. et al. Trophic downgrading of planet earth. Science, v. 333, n. 6040, p. 301–306, 2011.
FEARNSIDE, P. M. The Deforestation of the Brazilian Amazon: 9 – Roads. Blog Amazônia Real. 2020.
FINLAYSON, M. et al. Millennium Ecosystem Assessment Ecosystems and human well-being: wetlands and water synthesis. Washington, DC.: World Resources Institute, v. 1, 2005.
GARCÍA-NAVAS, V. et al. Temporal homogenization of functional and beta diversity in bird communities of the Swiss Alps. Diversity and Distributions, v. 26, n. 8, p. 900–911, 2020.
GUIMARÃES, D.; GARRASTAZU, M.; HIGUCHI, N. National System of Permanent Plots: proposal for a methodological model. Documentos – EMBRAPA Florestas (Brazil)., p. 67, 2005.
HADDAD, N. M. et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Advancement of science, v. 1, n. March, p. 1–10, 2015.
HARRIS, D. J. et al. Large trees in tropical rain forests require big plots. Plants, People, Planet, v. 3, n. 3, p. 282–294, 2021.
INAGUE, G. M.; ZWIENER, V. P.; MARQUES, M. C. M. Climate change threatens the woody plant taxonomic and functional diversities of the Restinga vegetation in Brazil. Perspectives in Ecology and Conservation, v. 19, n. 1, p. 53–60, 1 jan. 2021.
INEP. Notas estatísticas do censo da educação do ensino superior 2020. Censo da Educação Superior, Brasília MEC, 2022.
JAN, N.; LUDO, V. E. Software survey: VOSviewer, a computer program for bibliometric mapping. Cientometrics, v. 84, p. 523–538, 2010.
KOLAR, C. S.; LODGE, D. M. Progress in invasion biology: predicting invaders. Trends in Ecology & Evolution, v. 16, n. 4, p. 199–204, 2001.
KRAMER, J. M. F.; ZWIENER, V. P.; MÜLLER, S. C. Biotic homogenization and differentiation of plant communities in tropical and subtropical forests. Conservation Biology, n. October 2022, p. 1–13, 2022.
LAMBDON, P. W.; LLORET, F.; HULME, P. E. Do non-native species invasions lead to biotic homogenization at small scales? The similarity and functional diversity of habitats compared for alien and native components of Mediterranean floras. Diversity and Distributions, v. 14, n. 5, p. 774–785, 2008.
LAURANCE, W. F. et al. Rain forest fragmentation and the proliferation of successional trees. Ecology, v. 87, n. 2, p. 469–482, 2006.
LEWIS, S. L.; EDWARDS, D. P.; GALBRAITH, D. Increasing human dominance of tropical forests. Science, v. 349, n. 6250, p. 827–832, 2015.
LOBO, D. et al. Forest fragmentation drives Atlantic forest of northeastern Brazil to biotic homogenization. DIVERSITY AND DISTRIBUTIONS, v. 17, n. 2, p. 287–296, 2011.
MARTINEZ, O. J. A. Invasion by native tree species prevents biotic homogenization in novel forests of Puerto Rico. PLANT ECOLOGY, p. 1–16, 2010.
MAXWELL, S. L. et al. Biodiversity: The ravages of guns, nets and bulldozers. Nature 2016 536:7615, v. 536, n. 7615, p. 143–145, 10 ago. 2016.
MCKINNEY, M. L. On predicting biotic homogenization: Species-area patterns in marine biota. Global Ecology and Biogeography Letters, v. 7, n. 4, p. 297–301, 1998.
MCKINNEY, M. L.; LOCKWOOD, J. L. Biotic homogenization: a few winners replacing many losers in the next mass extinction. TREE, v. 5347, n. Table 1, p. 450–453, 1999.
MELLO, K. DE et al. Multiscale land use impacts on water quality: Assessment, planning, and future perspectives in Brazil. Journal of Environmental Management, v. 270, n. June, p. 110879, 2020.
MEYER, J.-Y. et al. The importance of novel and hybrid habitats for plant conservation on islands: a case study from Moorea (South Pacific). BIODIVERSITY AND CONSERVATION, p. 83–101, 2015.
MITTERMEIER, R. A. et al. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions. 1. ed. Mexico: Conservation International; CEMEX, 392p. p. 2004.
OUZZANI, M. et al. Rayyan—a web and mobile app for systematic reviews. Systematic Reviews, v. 5, n. 1, p. 210, 5 dez. 2016.
O’DEA, R. E. et al. Preferred reporting items for systematic reviews and meta-analyses in ecology and evolutionary biology: a PRISMA extension. Biological Reviews, v. 96, n. 5, p. 1695–1722, 2021.
OLDEN, J. D.; POFF, N. L. Ecological processes driving biotic homogenization: testing a mechanistic model using fish faunas. Ecology, v. 85, n. 7, p. 1867–1875, 2004.
OLDEN, J. D.; ROONEY, T. P. On defining and quantifying biotic homogenization. Global Ecology and Biogeography, v. 15, n. 2, p. 113–120, 2006.
OLIVEIRA, F. M. P. et al. Effects of increasing aridity and chronic anthropogenic disturbance on seed dispersal by ants in Brazilian Caatinga. Journal of Animal Ecology, v. 88, n. 6, p. 870–880, 2019.
PAIN, A. et al. What Is Secondary about Secondary Tropical Forest? Rethinking Forest Landscapes. Human Ecology, v. 49, n. 3, p. 239–247, 2021.
PARK, D. S.; RAZAFINDRATSIMA, O. H. Anthropogenic threats can have cascading homogenizing effects on the phylogenetic and functional diversity of tropical ecosystems. ECOGRAPHY, v. 42, n. 1, SI, p. 148–161, 2019.
PENN, J. L.; DEUTSCH, C. Avoiding ocean mass extinction from climate warming. Science, v. 376, n. 6592, p. 524–526, 2022.
PYSEK, P.; RICHARDSON, D. M. The biogeography of naturalization in alien plants. Jornal of biogeography, v. 33, p. 2040–2050, 2006.
RAHEL, F. J. Homogenization of fish faunas across the United States. Science, v. 288, n. 5467, p. 854–856, 2000.
RAHEL, F. J. Homogenization of freshwater faunas. Annual Review of Ecology and Systematics, v. 33, p. 291–315, 2002.
REES, M. et al. Long-Term Studies of Vegetation Dynamics. Science, v. 293, n. 5530, p. 650–655, 2001.
SANAPHRE-VILLANUEVA, L. et al. Patterns of plant functional variation and specialization along secondary succession and topography in a tropical dry forest Patterns of plant functional variation and specialization along secondary succession and topography in a tropical dry forest. Environmental research Letters, v. 12, n. 5, 2017.
SFAIR, J. C. et al. Taxonomic and functional divergence of tree assemblages in a fragmented tropical forest. ECOLOGICAL APPLICATIONS, v. 26, n. 6, p. 1816–1826, 2016.
SILVA, J. M. C. DA; TABARELLI, M. Tree species impoverishment and the future ¯ ora of the Atlantic forest of northeast Brazil. Nature, v. 404, n. March, p. 72–74, 2000.
SOLAR, R. R. DE C. et al. How pervasive is biotic homogenization in human-modified tropical forest landscapes? Ecology Letters, v. 18, n. 10, p. 1108–1118, 2015.
TOBY KIERS, E. et al. Mutualisms in a changing world: an evolutionary perspective. Ecology Letters, v. 13, n. 12, p. 1459–1474, 2010.
TYLIANAKIS, J. M.; TSCHARNTKE, T.; LEWIS, O. T. Habitat modification alters the structure of tropical host–parasitoid food webs. Nature, v. 445, n. 7124, p. 202–205, 2007.
URIBE, S. V; GARCÍA, N.; ESTADES, C. F. Effect of Land Use History on Biodiversity of Pine Plantations. Frontiers in Ecology and Evolution, v. 9, 2021.
VAN ECK, N. J.; WALTMAN, L. Visualizing Bibliometric Networks. Em: Measuring Scholarly Impact. [s.l.] Springer International Publishing, p. 285–320, 2014.
VAN ECK, N.; WALTMAN, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics, v. 84, n. 2, p. 523–538, 2010.
ZHANG, Y. M. et al. Biotic homogenization increases with human intervention: implications for mangrove wetland restoration. ECOGRAPHY, v. 2022, n. 4, 2022.
ZUPIC, I.; ČATER, T. Bibliometric Methods in Management and Organization. Organizational Research Methods, v. 18, n. 3, p. 429–472, 2015.
ZWIENER, V. P. Climate change as a driver of biotic homogenization of woody plants in the Atlantic Forest. Global Ecology and Biogeography, v. 27, p. 298–309, 2018.
[1] Ph.D. Candidate in the Graduate Program in Ecology and Natural Resources. UENF – Universidade Estadual do Norte Fluminense Darcy Riberio – Center for Biosciences and Biotechnology (CBB). Avenida Alberto Lamego 2000 Parque Califórnia 28013620 – Campos dos Goytacazes, RJ – Brazil. E-mail: [email protected]/; [email protected]. ORCID: 0000-0001-9700-796X. Currículo Lattes: https://lattes.cnpq.br/2743178685516399.
[2] Advisor. Associate Professor, Head of the Laboratory of Environmental Sciences (LCA). UENF – Universidade Estadual do Norte Fluminense Darcy Riberio – Center for Biosciences and Biotechnology (CBB). Avenida Alberto Lamego 2000 Parque Califórnia 28013620 – Campos dos Goytacazes, RJ – Brazil. E-mail: [email protected]. ORCID: 0000-0003-4492-3344. Currículo Lattes: http://lattes.cnpq.br/3704305950005564.
Submitted: May 26, 2023.
Approved: June 20, 2023.