TESE
PIRES, Ely [1], PIETROWISK, Vanda [2]
PIRES, Ely. PIETROWISK, Vanda. Dieta artificial para criação de Erinnyis ello, visando a produção em grande escala de baculovírus. Revista Científica Multidisciplinar Núcleo do Conhecimento. Ano 04, Ed. 10, Vol. 01, pp. 83-99. Outubro de 2019. ISSN: 2448-0959, Link de acesso: https://www.nucleodoconhecimento.com.br/agronomia/dieta-artificial
RESUMO
O experimento foi implantado com o intuito de formular uma dieta artificial para criação massal de Erinnyis ello L. (1578) (Lepidoptera: Sphingidae) para produção em grande escala do baclovírus EreLGV, como bioinseticida no controle do E. ello. O trabalho foi conduzido no laboratório de Ciências Biológicas da Unioeste, localizada no município de Cascavel–Pr. O delineamento utilizado foi o de delineamento inteiramente casualizado, no qual foram estabelecidos nove tratamentos, sendo denominados pelas dietas artificiais (D1, D2, D3, D4, D5, D6, D7, D8, D9) e folhas de mandioca da variedade baianinha como testemunha. As diferenças entre os tratamentos foram expressadas pelas quantidades e tipos de ingredientes inseridos em cada dieta. Foram realizadas 60 repetições, sendo cada uma delas representadas por uma lagarta. As lagartas inicialmente foram individualizadas em potes plásticos de 100 ml, e ao atingir o terceiro instar foram transferidas para potes de 1000 ml, permanecendo neste até a fase a adulta. Os parâmetros avaliados foram através das médias obtidas das: viabilidades larvais, pré-pupa, pupa, adultos e peso de pupas. O maior valor comparativo encontrado em todas as fases do inseto foi obtido através da dieta formulada (D6) seguida pelas dietas artificiais (D5 e D4). As dietas (D8 e D9) apresentaram as piores dietas, tendo 100% de mortalidades larvais. As demais dietas (D1, D2, D3, D7) foram as intermediárias em relação ao desenvolvimento de E. ello criadas em laboratório. Os resultados analisados demonstram que a dieta artificial formulada (D6) sobressaiu em relação ao desenvolvimento deste inseto, quando comparados as demais dietas avaliadas, sendo está a mais adaptada a criação massal de E. ello em meio artificial.
Palavra-chave: Nutrição de insetos, Manihot esculenta Crantz, Baculovírus erinnyis, alternativas de controle.
1. INTRODUÇÃO
O Erinnyis ello L. 1758 (Lepidoptera: Sphingidae), conhecido mais popularmente como mandarová-da-mandioca é considerado uma das principais pragas da cultura da mandioca (Manihot esculenta Crantz), devido seu alto potencial consumo foliar (FAZOLIN et al., 2007). Após o quarto instar chega a consumir até 75% de desfolha (BARRIGOSI et al., 2012) e segundo Farias (1991) durante período larval a média de área consumida atinge até 1.107 cm2.
Há evidências de que E. ello seja uma praga originária do Brasil, sendo constatada nos estados do Rio Grande do Sul e São Paulo no final do século XIX (FAZOLIN et al., 2007). Este inseto apresenta ampla capacidade migratória, tendo estudos comprovados que a distribuição geográfica ocorre desde a América do Sul até a América do Norte na fronteira com o Canadá Bellotti et al. (1983).
É uma praga esporádica, mas no Brasil pode-se observar incidência populacional em todos os meses do ano, variando de acordo com a temperatura elevada e o período chuvoso a determinada região (BELLOTTI et al., 1983; FARIAS, 2002).
A larva de E. ello pode se alimentar de mais de 35 diferentes espécies vegetais, abrangendo 10 famílias, das quais a mais importante é a Euphorbiaceae (BELLOTTI e ARIAS, 1982). Dentre as Euforbiaceas, três espécies são as principais hospedeiras: Manihot esculenta, Hevea brasiliensis (seringueira) SANTOS et al., 2014). Além de outras plantas de importância econômica como mamoeiro (Carica papaya), tomateiro (Lycopersicon esculentum), fumo (Nicotiana tabacum), algodoeiro (Gossypium herbaceum) e goiabeira (Psidium spp.) (CARVALHO, 1980; RITZINGER; SOUZA, 2000).
Dentre as diversas alternativas de controle do mandarová-da-mandioca destacam-se os usos do controle químico e biológico, contudo as estratégias de controle biológico apresentam-se cada vez mais promissoras, como é o caso do uso do Baculovírus (EreLGV).
O Baculovírus (EreLGV) é da família Baculoviridae que é uma das mais estudadas em relação controle de pragas da ordem lepidóptera (ARDISSON-ARAUJO, 2014). Trata-se de um vírus ovalado, com tamanho inferior a 0,5 micrômetros, apenas um virion por envelope e que o DNA das partículas virais denominada de EeGV, sendo este atuando especificamente ao controle do E. ello (SIHLER et al., 2014; ARAÚJO et al., 2014).
No Brasil, o vírus foi descoberto pela primeira vez na cidade de Cruzeiro do Sul – SC, no ano de 1986. Larvas coletadas em campo com possíveis sintomas de viroses foram encaminhadas a Empresa de Pesquisa Agropecuária de Santa Catarina (EPASC) para estudos e comprovação do agente biológico (ARAÚJO et al., 2014).
A multiplicação natural do vírus a campo apresenta-se controle ineficiente do Mandarová, devido à baixa multiplicação do vírus relacionado a alta incidência populacional e níveis de danos expressivos deste inseto a cultura da mandioca. Criação do mandarová em laboratório em dieta artificial como hospedeiro do Baculovírus (EreLGV) pode ser um dos principais sistemas de multiplicação do vírus no emprego de forma eficiente no controle do E. ello.
Ao se tratar de um controle altamente eficiente e ambientalmente seguro quando comparado ao controle químico e a necessidade de aplicação adicional de vírus (controle biológico clássico), tornou-se necessário desenvolvimentos de dietas artificiais adaptadas a criação massal do E. ello para produção industrial em grande escala do Baculovírus (EreLGV). Uma das principais dificuldades de produção de vírus entomapatogênicos pelo processo in vivo é a obtenção de grande quantidade de insetos (FINKLER, 2012).
As maiores eficiências de controles do mandarová-da-mandioca através do Baculovírus (EreLGV) ocorrem em larvas de até terceiro ínstar, equivalentes a 3 cm de comprimento e quando obter-se através de monitoramento 5 a 7 larvas pequenas por planta (MAIA; BAHIA, 2010; AGUIAR et al., 2010). Em experimento realizado por Maia; Bahia (2010) obtiveram 98% de mortalidades em larvas de terceiro instar.
A infecção e morte larval ocorre através da ingestão das partículas virais de Baculovírus (EreLGV). Ao atingir o mesêntero do intestino médio do inseto, a proteína que reveste o DNA do vírus é degradada e libera os virions (partícula infectiva) que invadem e multiplicam-se nas células do intestino, iniciando-se o processo de infecção e invadindo posteriormente a hemocele do inseto, multiplicando-se e ocasionando a sua morte (ALVES et al., 1998; ARAÚJO et al., 2014; ARORA, 2015).
Geralmente quatro dias após a ingestão das partículas virais a larva para de se alimentar, não ocasionando mais danos. Períodos que antecedem a morte, ou seja, aproximadamente seis dias após a infecção a larva migra para o dossel da planta, ficando dependurada no pecíolo da planta. Os sintomas observados nas larvas após a ingestão e multiplicações do vírus incluem descoloração da cutícula, perda de movimento e incapacidade de alimentação (ARAÚJO et al., 2014; ARORA, 2015).
Para Secchi (2002), o emprego do baculovírus oferece diversas vantagens ecológicas, devido apresentar controle específico a determinada espécie de inseto, além de oferecer segurança a outros organismos que atuam no controle de pragas. Em relação ao controle químico, o bioinseticida apresentam maior proteção ao meio ambiente, sendo alternativas mais sustentáveis e ecológicas.
2. MATERIAL E MÉTODOS
O experimento foi conduzido entre janeiro de 2017 a fevereiro de 2019 no Laboratório de Biotecnologia da Universidade Estadual do Oeste do Paraná-Unioeste, localizada no município de Cascavel-Pr, tendo as coordenadas geográficas de latitudes (24 56’ 26” S) e longitudes de (53 33’ 32” O) e 685 m de altitudes.
Para avaliar os desenvolvimentos biológicos de cada fase de E. ello, foram desenvolvidos nove tipos diferentes de dietas artificiais (D1, D2, D3, D4, D5, D6, D7, D8, D9), sendo as diferenças baseadas nas quantidades e tipos de ingredientes presente em cada uma das dietas avaliadas (Tabela 1).
Adultos de E. ello foram mantidos em gaiola com dimensão de 1,7×1,0x3,0 sendo esta construída no interior da casa de vegetação. Os adultos foram alimentados com solução de 10% de mel e água. No interior da gaiola foram colocados vasos de plantas com mandioca, sendo estas servindo para oviposição.
Com auxílio de pincel de cerdas macias, foram cuidadosamente coletados os ovos nas plantas de mandioca presentes na gaiola de criação, estes foram lavados com hipoclorito de sódio a uma concentração de 2% por 3 minutos e enxaguado em água destilada por três vezes, a fim de evitar contaminação, principalmente por vírus. Os ovos foram depositados sobre um papel filtro umedecidos com água para completarem seu desenvolvimento embrionário.
Após a eclosão, as larvas foram individualizadas em potes plásticos descartáveis de 100 mL contendo pedaços de dietas de cada respectiva dieta artificial. Permaneceram nestes potes até o 3º ínstar, quando foram então, em virtude do seu tamanho, transferidas para potes plásticos descartáveis de 1000 mL. Ao atingir o 5º instar foram acrescentados 5 cm de vermiculita ao fundo do pote simulando o solo, no qual será utilizado para o empupamento do inseto.
Quarenta e oito horas após a formação da pupa, estas foram pesadas com auxílio de balança de precisão e sexadas em microscópio estereoscópio. Estas foram novamente colocadas nos frascos com vermiculita até a emergências dos adultos.
O ensaio foi conduzido até início da fase adulta anotando-se mortalidade de cada ínstar, de pré-pupa e de pupa; duração dos ínstares e do período larval, de pré-pupa e pupa; peso de pupa e porcentagem de adultos deformados. A limpeza dos potes, avaliações e substituições da dieta foram realizadas diariamente.
Tabela 1 – Diferentes tipos de dietas e quantidades de ingredientes baseado a partir da dieta básica para criação do E. ello
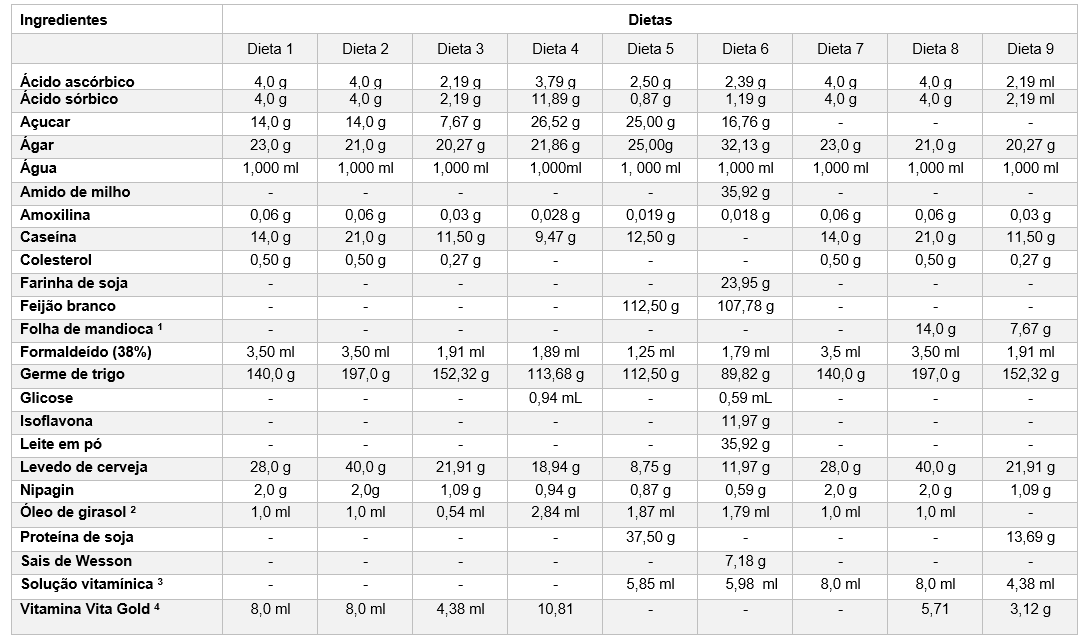
1 variedade baianinha, 2 substitui o Ácido. Linoleico, 3 Complexo vitamínico Vanderzant, 4 Vitamina A – 15,000,000,00 U.I.; Vitamina D3 – 4,000,000,00 U.I.; Vitamina E – 1,000,00 mg U.I.; Vitamina B1 – 4,000,00 mg U.I.; Vitamina B2 – 1,500,00 mg U.I.; Vitamina B6 – 20000,0 mg U.I.; Vitamina B12 – 4,800,00 mcg U.I.
Para obtenção das folhas de mandioca desidratada, presentes nas dietas 8 e 9, foram coletadas aleatoriamente folhas do terço apical, médio e basal da planta, variedade Baianinha. Estas foram desinfestadas em hipoclorito a 3%, durante 3 min e posteriormente colocadas em sacos de papel Cráft e levadas estufa de circulação forçada de ar (55 ºC) por até 72 horas, conforme Bueno et al. (2017). Após este procedimento as folhas foram maceradas com auxílio de um cadinho e pistilo até a formação de pó para posteriormente ser inserida nas dietas (D8 e D9).
O experimento foi realizado em delineamento inteiramente casualizado com 60 repetições, sendo que cada repetição foi constituída de uma larva individualizadas e 9 tratamentos, denominados pelas dietas artificiais (D1, D2, D3, D4, D5, D6, D7, D8, D9). Os dados coletados foram submetidos a análise de variância (ANOVA), as médias foram comparadas pelo teste de Tukey ao nível de 5%. O software utilizado para a análise estatística foi o Minitab 18.
3. RESULTADO E DISCUSSÃO
3.1 ENSAIO PARA AJUSTE DA DIETA BASE PARA DETERMINAÇÃO DE DIETA ARTIFICIAL PARA CRIAÇÃO DE E. ello.
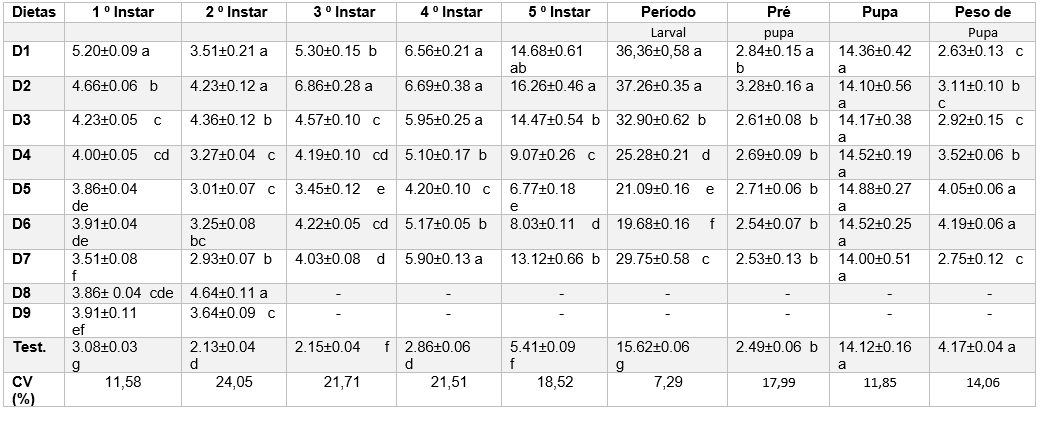
Observa-se que todas as dietas testadas alongaram os ínstares quando comparada às larvas alimentadas com folhas de mandioca. As dietas 5 e 6 foram as que, considerando o desenvolvimento larval apresentaram valores mais próximos à testemunha, embora diferindo estatisticamente destas (Tabela 2).
Devido ser as únicas dietas artificiais (D8 e D9) apresentarem em sua composição folhas desidratadas de mandioca e mortalidades larvais de 100% como demonstrado no final do 2º instar da tabela 2, acredita-se que as folhas possam apresentar fator antifagoestimulante ou outros compostos responsáveis pela mortalidade total deste inseto. Segundo Parra (2009) as plantas liberaram aleloquímicos que podem interferir no desenvolvimento do inseto. Trabalhos realizados (BAVARESCO et al., 2005), (CORRÊA, 2006) estão em acordo com este trabalho em que apresentou – se 100% de mortalidade antes da larva atingir o 3º instar de Hypocala andremona (Stoll, 1781) com folhas desidrata de caqui e Condylorrhiza vestigialis com folhas de Populus.
A duração entre os instares, apresentou-se grande variações, tendo a cada instar avaliados as larvas submetidas as dietas D5 e D6 apresentaram menores prolongamentos de dias entre as larvas submetidas as demais dietas. Apesar das variações de dias entre os instares larvais relacionados as diferentes dietas avaliadas, nota-se que ao 5º instar a D6 proporcional menor período total as larvas com 19.68, mesmo diferenciando estatisticamente, mas sendo este o próximo a da testemunha com 15.62 dias (Tabela 2).
Para a duração do período de pré-pupa observou-se que os maiores prolongamentos ocorreram nas dietas D1 com 2.84 e D2 com 3.28 dias e para as demais dietas não houve diferenças estatísticas (Tabela 2).
Não houve diferença estatística para duração do período de pupa entre as dietas avaliadas. Em relação a duração do período larval, observou-se que as dietas D1 36.36 e D2 37.26 dias não diferenciando estatisticamente entre si. Já para as demais dietas observa-se que houve diferenças estatísticas entre si, mas a que se aproximou-se mais da testemunha (folhas de mandioca) foi a dieta D6 com 19.68 dias (Tabela 2).
Para peso de pupa as dietas D5 e D6 tiveram valores similares aos da testemunha, indicando que, mesmo alongando o período larval, as larvas conseguiram ingerir a quantidade necessária de nutrientes. Segundo Barrigossi et al. (2012) as larvas de E. ello aumentam a taxa de consumo quando o nível nutricional do alimento não é adequado. As demais dietas apresentaram pupa com pesos inferiores aos das alimentadas com folhas de mandioca, demostrando que, mesmo alongando o período larval, no qual a larva teve mais tempo para ingerir a dieta, ao final houve carência de nutrientes, formando menos massa de pupa (Tabela 2)
Tabela 2 – Duração (dias) dos instares e fases de Erinniys ello (Lepidoptera:Sphingidae) criados em diferentes dietas artificiais(D1 a D9) em folhas de Mandioca da variedade Baianinha.
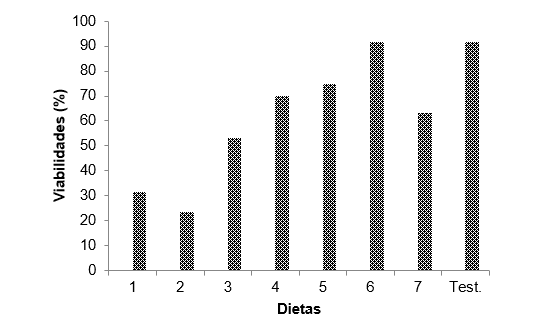
Segundo Singh (1983) e Parra (2012), para que uma dieta seja adequada a criação massal de um inseto, esta deve possibilitar viabilidade larval igual ou superior a 75,0%. Portanto através da figura 1, pode-se observar que as dietas D5 e D6 por apresentarem 75% e 91.7% respectivamente de viabilidade, são as únicas entre as demais apresentar aptas a criação massal em laboratório, conforme citado pelos autores.
Quanto as viabilidades larvais inferiores 75% (Figura 1) podem estar relacionados as diversas condições nutricionais, físicas e fagoestimulantes. Segundo (PARRA, 2002), os índices elevados de mortalidade larvais podem ser indicativos de inviabilidade da dieta. As deficiências quantitativas e qualitativas em uma dieta podem afetar diretamente a sobrevivência do inseto (PARRA, 1991).
Figura 1 – Porcentagem de viabilidade total de larvas de Erinnyis ello alimentadas com diferentes dietas artificiais e com folha de mandioca, variedade baianinha.
Em relação a viabilidade de pupa, obteve-se maiores sobrevivências na dieta artificial D5 com 97,8%, seguida pela D6 atingindo 96,4%. Viabilidades intermediárias ocorreram nas dietas D3, D4, D7 e testemunhas, com 91,2%, 92.7%, 93,8%, 94,54%, respectivamente. As dietas artificiais D1 e D2 apresentaram as menores viabilidades pupais sendo respectivamente 89.5%, 86,7% (Figura 2).
As sobrevivências de pupas apresentaram variações entre as dietas avaliadas. Não houve diferenças estatísticas entre as D5 e D6, que por sua vez se diferenciaram das demais dietas. Em relação às dietas intermediárias D4, D3, D7 e testemunha não diferenciaram entre si (Figura 2).
Os resultados demonstram alta viabilidade de adultos cujas larvas foram alimentadas com as diferentes dietas. Em nenhuma das dietas foram observadas inviabilidades inferiores a 80,0%. Nota-se que nenhuma das dietas da fase larval influenciou a viabilidade dos adultos, quando a larva conseguiu empupar. A dieta D3, por exemplo, considerada uma das piores dietas, quando avaliada a fase de larva, apresentou viabilidade superior (94,1%), enquanto que a melhor dieta (D6) teve 89,1%, juntamente com a testemunha, com 90,9% (Figura 2).
Figura 2 – viabilidades de pupas e adultos de Erinnyis ello (Lepidoptera: Sphingidae) alimentadas com diferentes dietas artificiais.
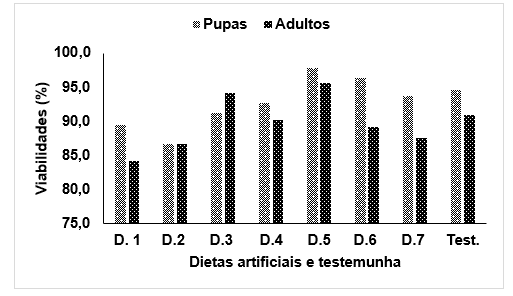
O desenvolvimento observado nos insetos submetidos às dietas D5 e D6 podem estar relacionados ao aumento de fagoestimulantes, sendo 20 g de sacarose (D5) e 14 g de sacarose mais 0,5 g de glicose na D6, sendo estas duas dietas com maiores quantidades de fagoestimulantes em relação às demais (Tabela 1). Os fagoestimulante podem ter propiciado maior estímulo alimentar e consequentemente maior consumo e desenvolvimento dos insetos submetidos a estas dietas. Diversos autores relatam a importância dos fagoestimulantes no consumo da dieta pelo inseto (PARRA, 1996; PARRA, 1998; PARRA, 2002; SOUZA et al., 2009).
A D6, além de apresentar o principal quesito mencionados por Singh (1983) e Parra (2012) que é de apresentar no mínimo de 75,0% de viabilidade para que seja considerada uma dieta adaptada a criação massal em laboratório, apresentou-se ainda de baixo custo, devido as eliminações do colesterol e caseína e as reduções do ágar e dos anticontaminantes que são ingredientes de custos elevados e presentes nas demais dietas avaliadas neste experimento. Segundo Parra (1998), o ágar é responsável por aumentar o custo de uma dieta artificial entre 60,0 a 70,0%.
Não houve a substituição do ágar, mas o amido de milho pode ter contribuído pelo aumento da resistência e consequentemente reduzindo a quantidade de ágar e o custo da dieta, sendo este ingrediente contendo unicamente na dieta artificial D6 (Tabela 1). O emprego do amido de milho como geleificante na substituição parcial do ágar em dietas para criação de Spodoptera frugiperda, apresentou redução de até 40.6% de custo (PERES et al., 2010).
As variações dos diversos parâmetros biológicos do mandarová-da-mandioca relacionados aos diferentes tipos de dietas avaliados demonstraram que os piores resultados foram observados nas dietas D1, D2, D3, D7, D8, D9) (Tabela 2), (Figuras 1 e 2).
Embora tenha se mostrado viável a criação do mandarová-da-mandioca, é prematuro afirmar que a D6 é viável, uma vez que não foi possível avaliar várias gerações criadas na dieta e também a fecundidade. Nesse sentido, Ribeiro (2017) considerou a adequação da dieta artificial para criação de Helicoverpa armigera após 12 gerações. Serão necessários que sejam realizadas avaliações durante várias gerações para confirmar a adequação de uma dieta artificial em relação a uma determinada espécie de inseto (COHEN, 2015).
4. CONCLUSÃO
De acordo com as condições desta pesquisa pode-se considerar que dieta artificial formulada D6 se apresentou-se ser a mais adaptada para a criação de E. ello em laboratório, demonstrando maiores viabilidades em todas as fases biológicas avaliadas neste experimento.
5. REFERÊNCIAS
AGUIAR, E. B.; LORENZI, J. O.; MONTEIRO, D. A.; BICUDO, S. J. Monitoramento do mandarová da mandioca (Erinnyis ello L. 1758) para o controle com baculovírus (Baculovírus erinnyis). Revista Trópica: Ciências Agrárias e Biológicas, Chapadinha, v. 4, n. 2, p. 55-59, 2010.
ALVES, S. B. Produção de vírus entomapatogênicos. In: ALVES, S. B. (Coord.). Controle microbiano de insetos. 2. ed. Piracicaba: FEALQ, p. 871-887, 1998.
ARAUJO, D. M. P. A.; MELO, F. L. de.; ANDRADE, M. de S.; SHLER, W.; BÁO, S. N.; RIBEIRO, B. M.; SOUZA, M. L. de. Genome sequence of Erinnyis ello granulovírus (EreLGV), a natural cassava hornworm pesticide and the first sequences sphingid-infecting betabaculovírus. Bmc Genomics. Brasilia, DF, Brasil, 2014.
ARDISSON-ARAUJO, D. M. P. A.; MELO, F. L. de.; ANDRADE, M. de S.; SHLER, W.; BÁO, S. N.; RIBEIRO, B. M.; SOUZA, M. L. de. Genome sequence of Erinnyis ello granulovírus (EreLGV), a natural cassava hornworm pesticide and the first sequences sphingid-infecting betabaculovírus. Bmc Genomics, v. 15, p. 853 – 864. 2014.
ARORA, R. Microbial control in insect pest manegament: Achievements and challengs. P. 97 -152 IN: SINGH, B.; ARORA, R.; GOSAL, S.S. (Eds). Biological and moleculares approaches in pest management. Ludhiana, India. 450 p. 2015. Disponível em:https://books.google.com.br/books?hl=ptBR&lr=&id=u6I4DwAAQBAJ&oi=fnd &pg=PA97&dq=betagranulovirus+Erinnyis+ello&ots=o4hfKzm-di&sig=jsvCxAqZ757 t0sjEw3AesaGmas4#v=onepage&q&f=false
BARRIGOSSI, J. A. F.; ZIMMERMANN, F. J. P.; LIMA, P. S. C. Consuption rates and performance of Erinnyis ello L. on four cassava varieties. Neotropical Entomology, v. 31, n. 3, p. 429-433, 2012.
BAVARESCO, A.; GARCIA, M. S.; BOTTON, M.; NONDILLO, A. Avaliação de dietas artificiais para criação de Hypocala andremona (STOLL, 1781) (Lepidoptera: Noctuidae). Ciência Agroveterinárias, Lages, v. 4, n. 2, p. 94-100, 2005.
BELLOTTI, A. C.; ARIAS, V. B. El control de Erinnyis ello (L) (gusano cachón de la yuca). Cali: CIAT, 1982. 24p.
BELLOTTI, A. C.; REYES, J. A.; VARGAS, O.; ARIAS V., B.; GUERRERO J. M. Descripción de las plagas que atacan la yuca (Manihot esculenta Crantz) y características de sus daños. Cali: CIAT, 1983. 51p.
BUENO, A. V. I.; JOBIM, C. C.; RIBEIRO, M. G.; OLIVEIRA, J. P. Método de obtenção de matéria seca e composição química de volumosos. Ciência Animal, Goiânia, v.18, p. 1-8, 2017.
CARVALHO, C.F. Aspectos Biológicos, técnicas para obtenção de ovos em condições de laboratório e avaliação de danos de Erinnyis ello (L., 1758) (Lepidoptera: Sphingidae) em mandioca em condições de campo. 1980. 87p. Dissertação. ESALQ/USP.
COHEN, A. C. Insect diests. Sciense and technology. Boca Ratón: CRC Press, 2015. p. 1-164.
CORRÊA, F. A. S. F. Criação em laboratório de Condylorhiza vestigialis (Guenné 1824) (Lepidoptera: Crambidae) com diferentes dietas artificiais. 2006. 82 p. Dissertação (Mestrado em Ciências Florestais) – Universidade Federal do Paraná, Curitiba, 2006.
FARIAS, A. R. N. Insetos e ácaros pragas associadas a cultura da mandioca no Brasil e meios de controles. Cruz das Almas, BA: Embrapa – CNPMF, 1991. 47 p. (Circular Técnica, 14).
FARIAS, A. R. N. Pragas da Mandioca. In: OTSUBO, A. A.; MERCANTE, F. M.; MARTINS, C. S. (Eds). Aspectos do Cultivo da Mandioca em Mato Grosso do Sul. Dourados: Embrapa Agropecuária Oeste; Campo Grande: UNIDERP, 2002. p. 169-190.
FAZOLIN, M. et al. Manejo integrado do mandarova-da-mandioca Erinnyis ello (L.) (Lepidotera: Sphingidae): conceito e experiência na região do Vale do Rio Juruá, Acre. Rio Branco: Embrapa Acre, 2007. 45 p. (Documentos, 107).
FINKLER, C. L. L. Controle de inseto: Uma breve revisão. Anais da Academia Pernambucana de Ciência Agronômica, Recife, vols. 8 e 9, p. 169-189, 2012.
MAIA, V. B.; BAHIA, J. J. S. Manejo integrado do mandarová (Erinnyis ello L.) em mandioca (Manihot esculenta crantz) na região sul da Bahia. Ministério da Agricultura, Pecuária e Abastecimento – Mapa. p. 16, Bahia, 2010.
PARA, J. R. P. Consumo e utilização de alimentos por insetos. In: PANIZZI, A. R.; PARRA, J. R. P. (Eds.). Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo: Manolle, 1991. p. 9-65.
PARRA, J. R. P. Criação de insetos para estudos com patógenos. In: ALVES, S. B. (Coord.). Controle microbiano de insetos. 2. ed. Piracicaba: FEALQ, 1998. p. 1163.
PARRA, J. R. P. Criação massal de inimigos naturais. In: PARRA, J. R. P. et al. (Eds.). Controle biológico no Brasil. São Paulo: Manole, 2002. p. 143-164.
PARRA, J. R. P. Técnicas de criação de insetos para programas de controle biológicos. 4. ed. Piracicaba: FEALQ, 1996. 137p.
PARRA, J. R. P. The evolution of artificial diets and their interaction in science and technology. In PANIZZI, A. R.; PARRA, J. R. P (Eds) Insect biocology and nutrition for integraínstar pest management. Boca Ratón: CRC Press, 2012. p. 51-92
PARRA, J. R. P.; PANIZZI, A. R.; HADDAD, M. L. Nutricional indices for measuning food intake and utilization. In: PANIZZI, A. R.; PARRA, J. R. P. (Eds) Insect bioecology and nutrition for integraínstar pest management. Boca Ratón: CRC Press, 2012. P. 13-49.
PARRA, J. R. P. A evolução das dietas e suas interações em ciência e tecnologia. 3º. capitulo. Brasilia, DF: EMBRPA, 2009. 84p.
PERES, E. C.; FUGA, C. A. G.; GONSALVES, D. C. CUNHA, W. V. Avaliação de elementos geleificantes de baixo custo para substituição do ágar usado na elaboração de dieta artificial de Spodoptera frugiperda (Lepidoptera: noctuidae). Revista do Núcleo Interdisciplinar de Pesquisa e Extensão do Unipam. Ed. 7ª, v. 1, p. 194-2001, Patos de Minas, 2010.
RIBEIRO, Z. A. Dieta artificial e metodologia de criação massal para o bem-estar de Helicoverpa armigera. 2017.122 f. Tese (Doutorado em entomologia) Faculdade de Ciência Agrária – Unespe, Jabotical, 2017.
RITZINGER, C. H. S. P.; SOUZA, J. da S. Mamão fitossanidade. Embrapa comunicação para transferência de tecnologia. P35, Cruz da Almas – BA, 2000.
SANTOS, R. S.; TAVARES, M. T.; SUTIL, W. T.; VASCONCELOS, A. S.; AZEVEDO, T. S.; DIOGO, B. S. Parasitismos de Brachymeria annulata (Fabricius) (Hymenoptera: Chalcildidaea) em Erinnyis ello (L.) (Lepidoptera: Sphingidae). Rio Branco, Ac, 2014.
SECCHI, V. A. Baculovírus, mais do que uma grande descoberta: uma revolucionária alternativa aos agrotóxicos. Agroecologia e Desenvolvimento Rural Sustentável, 2002.
SIHLER, W.; SANCHES, M. M.; FALCÃO, R.; FANZOLIN, M.; ESTRELA, J. L. V.; SOUZA, M. L. Carcaterização de Erinnyis ello granulovírus isolados de populações de Mandarová da Mandioca em Cruzeiro do Sul, Acre. XXV Congresso Brasileiro de Entomologia. Goiania-GO, 2014.
SINGH, P. A. General Purpose laboratory diet mixture for rearing insects. Insect Science and its Application, Oxford, v. 4, n. 4, p. 357-362, 1983.
SOUZA, D. M.; LÔBO, A. P.; BADJI, C. A. Efeito de fagoestimulantes no consumo de folhas de meloeiro por Diaphania hyalinata. In: Jornada de Ensino, Pesquisa e Extensão. n. 9. Anais UFRPE. Recife, 2009.
[1] Biólogo e Engenheiro Agrônomo. Doutorado na área de Produção Vegetal com ênfase em Entomologia; Mestrado na área de Produção Vegetal com ênfase em Nematologia; Graduações: Agronomia e Ciências Biológicas.
[2] Bióloga. Professora da disciplina de Entomologia da Universidade Estadual do Oeste do Paraná- Unioeste – Campus de Marechal Cândido Rondon – Pr.
Enviado: Maio, 2019.
Aprovado: Outubro, 2019.