ОБЗОР СТАТЬИ
SILVA, Izael Oliveira [1], AMORIM, Edna Peixoto da Rocha [2], NASCIMENTO JUNIOR, Nelson Augusto [3], PEIXINHO, Georgia Souza [4], CARNAÚBA, Juliana Paiva [5], NETO, Vicente Ferreira de Araujo [6]
SILVA, Izael Oliveira. Et al. Trichoderma spp антагонизм к фитопатогенам, вызывающим корневую гниль маниоки в алагоа. Revista Científica Multidisciplinar Núcleo do Conhecimento. 05-й год, Эд. 06, г. 12, стр. 149-177. Июнь 2020 года. ISSN: 2448-0959, Ссылка на доступ: https://www.nucleodoconhecimento.com.br/агрономия-ru/trichoderma-spp-антагонизм, DOI: 10.32749/nucleodoconhecimento.com.br/ru/55553
РЕЗЮМЕ
Некоторые из них фитопатогены, которые ограничивают производство маниока (Manihot esculata Crants) время гнили корней, к которым они женятся наибольшую потерю урожая достижения 70%. Эта культура представляет большой интерес для северо-восточного региона Бразилии, обеспечивая большие потери. Трудности в управлении направлены на поиск альтернатив, среди которых биоконтроллеры показали значительные результаты. Этот обзор литературы направлен на сбор информации, связанной с гнилью корня маниоки CV Rosinha, основные фитопатогены и антагонистической деятельности Trichoderma в борьбе с болезнями в штате Алагоас. Библиографическое исследование было проведено в базах данных Scielo, Google Acadêmico, портале Capes (тезисы и диссертации), журналах и книгах. Для получения информации о: корневая гниль таблицы маниока, Trichoderma spp., как биоконтроль агента фитопатогенов.
Ключевые слова: туберозный корень, биологический контроль, фитопатология.
1. ВВЕДЕНИЕ
Благодаря своей питательной ценности, Manihot esculenta Crantz выделяется в Бразилии и в мире со значительным экономическим и социальным вкладом из-за своей универсальности, так как он используется как в животных и человеческих продуктов питания и широко используется в промышленности. Урожай оказывает значительную поддержку населению с низкой покупательной способностью, поскольку помимо того, что он является питательным источником, он также является источником дохода в семейном сельском хозяйстве (NOTARO et al. 2013).
В мире потребление маниоки и ее производных находится на шестой позиции в рейтинге, где по данным FAO (2012) можно найти соевые бобы, пшеницу, рис, кукурузу и картофель. Бразилия является четвертым по величине производителем в мире, с оценочным урожаем за 2016 год в 25 миллионов тонн (IBGE, 2015).
Из большой универсальности, урожай полностью используется, а не только воздушная часть (листья и стебли), но и корень. Листья имеют их использование в человеческой (дополнение) и животных (раздавленных) пищи. Стебли, в кормах для животных, в виде силосов и сена и до сих пор в натуре. Корень, в продуктах питания человека, животных, а также промышленности, в основном мука, крахмал, биоразлагаемые пластмассы, текстильные изделия и биотопливо (CONAB, 2013).
Культура Кассавы сталкивается со многими проблемами, в том числе с фитосанитарными проблемами (NASCIMENTO JÚNIOR, 2015). Несколько заболеваний уже сообщалось в культуре, среди них, корневая гниль, чьи агенты могут быть Lasiodiplodia sp., Pythium sp., Rosellinia sp., Fusarium sp., Neoscytalidium lignicola, Phytophthora drechsleri Такер однако, последние три фитопатогена наиболее часто участвуют в корневой гнили в штате Алагоас (Рисунок 1) Информация о гнили корней маниоки по-прежнему зарождается и имеет первостепенное значение для того, чтобы служить инструментом при принятии решений о наилучшей стратегии использования (MASSOLA JR; BEDENDO, 2005; МUNIZ et al., 2006; NOTARO et al., 2013; SILVA, 2013; FUKUDA; OTSUBO, 2015).
Среди микроорганизмов, вызывающих корень маниоки гнили P. drechsleri и F. solani, они являются наиболее важными из-за масштабов и тяжести, вызывая значительные потери в производстве. В плотных и подкисленных почвах появление Fusarium sp., является более частым, и одна из характеристик этого патогена заключается в том, чтобы вызвать повреждение стебля вблизи почвы инфекцией ксилемических сосудов и, следовательно, вмешиваться в циркуляцию сока, тем самым вызывая косвенную гниль корня в любой причине развития растений. В отличие от Phytophthora, симптомы которой провоцируются в корнях коричневого цвета и мягкой консистенции гнили, которые источают жидкость с неприятным запахом, уже гнили Fusarium sp., без видимых нарушений в тканях с признаками обезвоживания и мумифицированного появления сухой гнили (MOURA; SILVA, 1997; POLTRONIERI et al., 2001; SILVA; TEIXEIRAS, 2012).
В качестве защитных и корневых мер по борьбе с гнилью используется физическое и химическое управление почвой, однако было положительно поддержано использование терпимых сортов, связанных с культурными практиками, такими, как севооборот и система культивирования. Nascimento Júnior (2015) подчеркивает, что принципиально важно знать эдафо-климатические условия региона культивирования и использовать соответствующие методы, которые могут быть приняты фермером, для обеспечения экономической отдачи в рамках устойчивой сельскохозяйственной системы. Альтернативный контроль фитопатогенов с антагонистами, особенно видами Trichoderma, уже изучен, в связи с мировым интересом к использованию продуктов, которые менее агрессивны к окружающей среде (SOUSA et al., 2012; SILVA et al., 2014). Для Robbs (1992) и Barbosa; Meza (2009) конкурс на питание и антибиоз, являются механизмами наиболее часто используются некоторыми видами Trichoderma, поэтому считается отличным агентом биоконтроля.
С учетом вышесказанного цель настоящей работы заключалась в проведении библиографического обзора основных фитопатогенов и потенциала антагонизма Trichoderma, связанного с гнилью корнеов маниоки в штате Алагоас.
2. МЕТОДОЛОГИЯ
Настоящее исследование состоит из обзора литературы, проведенного в период с декабря 2014 года по март 2016 года, в котором была проведена консультация по научным статьям, отобранным с помощью поиска в базе данных Google acadêmico, LILACS, MEDLINE, SciELO, портала Capes (диссертаций и диссертаций), журналов и книг. Из многочисленных связанных с этим статей 84 из них были использованы. В качестве дескрипторов мы использовали: корневую гниль, биологический контроль фитопатогенов, фитосанитарность.
3. ОБЗОР ЛИТЕРАТУРЫ
3.1 КУЛЬТИВИРОВАНИЕ МАНИОКИ
Кассава, культурой, представляющий большой интерес для бразильского северо-востока, особенно для штата Алагоас, является многолетним кустарником растение, принадлежащее к семье Euphorbiaceous, богатое крахмалом, используемое в продуктах питания для человека и животных или в качестве сырья для различных отраслей промышленности (FRAIRE FILHO; BAHIA, 2015).
Nascimento Júnior (2015), подчеркивает преимущества этой культуры, из-за его деревенскости, низкой стоимости, адаптации по отношению к климату, почве и его питательные характеристики, которые связаны с большим накоплением крахмала в его корнях, а также белков и аминокислот в листьях. Это растение, возникшее в тропических регионах южной Америки, но которое имело в качестве мест его одомашнивания юго-восточной Азии, Африки и тропической Америки (NOTARO et al., 2013; SILVA, 2013; FRAIRE FILHO; BAHIA, 2015).
В 2013 году во всем мире было произведено 281 718 000 тонн маниоки, при этом Бразилия занимает четвертое место в мире с 23 977 757 тоннами после Нигерии, Индонезии и Таиланда. Северные и северо-восточные регионы Бразилии имеют большой объем производства и культивируемых площадей находится на юге, юго-востоке и Среднем Западе, те, с самым высоким сельскохозяйственным урожаем. Штат Пара является крупнейшим национальным производителем корня маниоки, за которым следуют Парана, Мараньян и Баия (IBGE, 2015).
В Алагоасе регионы, которые выделяются как производители маниоки, являются дикими, с 70% от общего объема производства государства (Arapiraca, Сан-Себастьян и Кампо-Алегри), лесной зоны и северного побережья, будучи первым регионом, один с наибольшим числом производителей, ответственных за 13,36 t.ha-¹ и 240,448 тонн, занимая второе место по производительности по сравнению с другими государствами северо-востока (CONAB 2013).
Notaro (2012) подчеркнул, что эта культура по-прежнему является семейным фермерством и развивается в рудиментарном ключе, а основными средствами распространения является вегетативность и до сих пор выращиваются сорта низкого генетического качества. Его полезность связана с содержанием гидроциановой кислоты (HCN), присутствуют в листьях и корнях, что классифицирует его как кроткий или сладкий(<180 mg kg-¹ de HCN), также известный как “macaxeira”, используемый для потребления в свежем виде; промежуточные (180-300 mg kg-¹) и горькие или дикие (> 300 mg kg-¹) последние два используются для производства кормов для животных, крахмала, муки и биотоплива (OLIVEIRA et al., 2012).
Наибольшая полезность маниоки в пищевой промышленности, в основном в производстве крахмала, предназначенного для модифицированного рынка крахмала, который в дополнение к обслуживанию региональной кухни в хлеб, тапиоки, beijus, жевательный торт и ног сорванец; является одним из продуктов, полученных от завода, который использует больше труда и добавляет высокую ценность в переработанном продукте, в соответствии с Nascimento Júnior (2015). Что касается использования для добычи ферментированных продуктов в биотопливной промышленности, маниока была показана как хорошая альтернатива в производстве этанола, так как он представил более низкую стоимость энергии в производстве, сравнивая сахарный тростник и кукурузу, в соответствии с Salla et al., (2010).
Согласно Alves (2006), фенология маниоки может варьироваться от 6 до 24 месяцев культивирования, но коммерчески это от 12 до 18 месяцев для обработки муки и крахмала и от 8 до 10 месяцев для стола. Среди сортов, посаженных в Алагоасе, розина выделяется, чей цикл варьируется от 9 до 10 месяцев в суше, и может быть ожидать в течение 7 до 8 месяцев в условиях орошаемой системы. Этот сорт может достигать трех метров в высоту, поддерживает завод стенд в конце урожая с хорошим производством ветвей и производительности в 21 t ha-¹ (DINIZ et al., 2009).
Nascimento Júnior (2015), сообщает в своей работе, что даже при деревенскости урожая необходимо внимательно относиться к адафо-климатических условий, заявив, что маниока производит хорошо в плодородных почвах, с рН между 5,5 до 7,0 с песчаным или средним текстурным классом и хороший естественный дренаж. Идеальное климатическое состояние находится в температурном диапазоне от 20°C до 27°C градусов по Цельсию и может быть установлено в средах с температурой от 16°C до 38°C градусов по Цельсию; с осадками от 1000 до 1500 mm в год-1, в полузасушливых районах от 500 до 700 mm в год-1; высоты от 600 до 800 метров над уровнем моря и световой период 12 часов день-1 (SILVA; ANDRADE, 2011; SILVA, 2013).
3.2 ЗАБОЛЕВАНИЯ В УРОЖАЕ МАНИОКА
Несколько факторов могут ограничить производство маниоки, такие, как появление насекомых-вредителей и вызывающих болезни фитопатогенов, что ухудшает качество и развитие урожая в Бразилии и во всем мире. По словам Nascimento Júnior (2015), маниока подвержена воздействию нескольких фитопатогенов, что приводит к серьезным экономическим потерям в производстве.
Среди заболеваний, имеющих важное значение выделяются причины поражения листьев, таких как коричневое пятно (Cercosporidium henningsii) белое пятно (Phaeoramularia manihotis), горящие листья (Cercospora vicosae), черное пятно (C. manihobaea) и антракноз (Colletotrichum gloeosporioides f.sp. manihotis), которые ограничивают фотоассимилаты (SILVA; ANDRADE, 2011; MORAIS et al., 2013; OLIVEIRA et al., 2013). И корневая гниль, вызванная грибами и oomicetes, таких как Lasiodiplodia sp., Pythium sp., Rosellinia sp., Fusarium sp., Neoscytalidium lignicola, Phytophthora drechsleri Tucker однако, последние три фитопатогена являются наиболее часто участвующими в корневой гнили в штате Алагоас (Рисунок 1) Информация о гнили корней маниоки по-прежнему зарождается и имеет первостепенное значение для того, чтобы служить инструментом при принятии решений о наилучшей стратегии использования (MASSOLA JR; BEDENDO, 2005; MUNIZ et al., 2006; NOTARO et al., 2013; SILVA, 2013; FUKUDA; OTSUBO, 2015).
Notaro et al., (2013) и Silva (2013) предупреждают о высоких социально-экономических последствиях, вызванных гнилью корнеов маниоки в штатах Мараньян, Пара, Алагоас и Пернамбуку, так как это приводит к значительным потерям в производительности, в дополнение к тому, чтобы сделать районы нежизнеспособными для посадки на протяжении всего цикла сельскохозяйственных культур. По словам Nascimento Júnior (2015), будучи Omyceto Phytophthora sp и гриб Fusarium sp., являются основными патогенами культуры на северо-востоке и вызывают потери в диапазоне от 30 до 70%. Автор также утверждает, что при благоприятных условиях болезнь может вызвать 100% потерю.
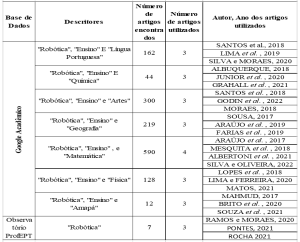
Рисунок 1. Аспект колоний и репродуктивных структур фитопатогенов, вызывающих маниоку гнили изолятов в штате Алагоас: Fusarium sp. (A1-A6), Neoscytalidium sp. (B1-B6) и Phytophthora sp. (C1-C6). Колония Fusarium sp. (A1); конидия (A2); конидиогенные клетки (A3-A6). Neoscytalidium sp. колония (B1); Конидия (B2-B6). Phytophthora sp колония (C1); Споманджио (C2); Хламидоспоры (C3 и C4); Споманджиос (C5) и Sporantia в воспроизводстве (C6).
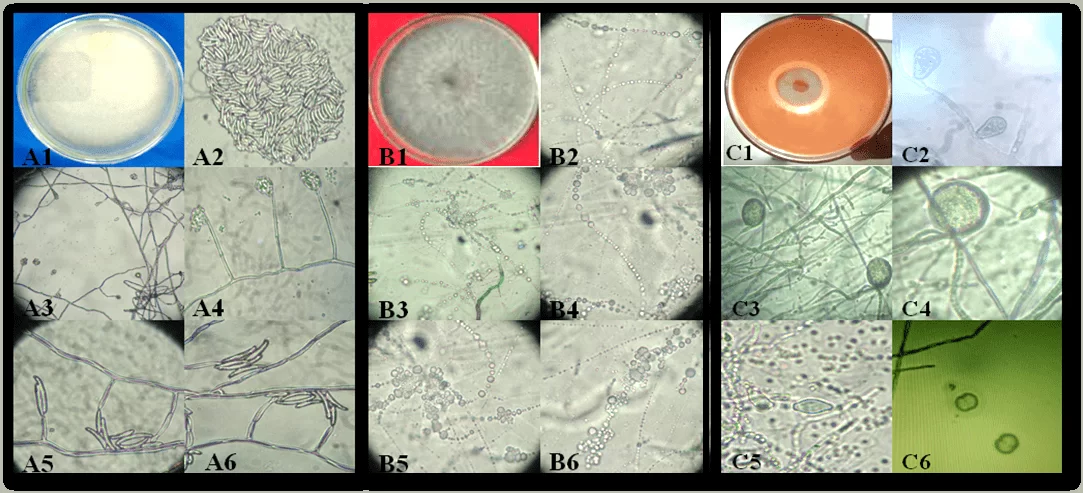
Гниль, вызванная Fusarium sp., происходит в молодых и взрослых растений, с симптомами внезапного увядания и тяжелой дефолиации, которые могут быть потенцирован в районах clayey и плохо осушенных почвы (SILVA; TEXEIRA, 2012). Этот факт уже был сообщен Gomes; Leal (2003), который при работе с маниокой в штатах Сержипе, Баия и Алагоас, утверждает, что корневая гниль является наиболее ограничивающей болезнью культуры в северо-восточном регионе, особенно когда она имплантируется в районы, образованные уплотненной почвой.
Фитопатогены, вызывающие гниль, имеют явные симптомы. Авторы сообщают, что Phytophthora sp. обычно атакует взрослые растения, вызывая «мягкую» гниль в корнях, с очень сильным запахом, похожим на разлагающееся органическое вещество и серовую окраску. Появление признаков более часто и видно в корнях, но Есть сообщения о существовании молодых людей, представляющих симптомы в основании стеблей новых близнецов растений, вызывая увядание и полную смерть. В случае сухой гнили, вызванной Fusarium sp., прямое повреждение корней редко происходит, но проблема усугубляется на любом этапе развития завода. (GOMES; LEAL, 2003; SILVA; TEXEIRA, 2012; NASCIMENTO JÚNIOR, 2015).
3.3 ФИТОПАТОГЕН ФУЗАРИУМ
Род Fusarium был описан в 1809 году Link. Представители этого рода принадлежат к царству Грибы, филум Ascomycota, классы Eauscomycetes и Pyrenomycetes, заказать Hypocreales, представляя 65 видов, 55 сортов и 22 специальных форм, в соответствии с Wollenweber; Reinking (1935)1 цитирует Ventura (1999). Это один из важнейших фитопатогенов в мире, с широким географическим распространением, распределенным в почве и в сотрудничестве с растениями. Наличие космополитических и других видов с появлением ограничивается определенными средами (VENTURA, 1999). Большинство видов являются сапробическими, но есть представители комплекса Fusarium, которые производят микотоксины, которые могут повлиять на здоровье человека и животных (URBEN et al. 2009). Seifert (2006); Querales (2010) сообщает, что род Fusarium представляет собой сложную группу, растущую по количеству видов и порождающий таксономические споры для фитопатологов.
Milanesi (2009), комментирует то Fusarium sp., может произойти специально в местах тропического и субтропического климата, и имеет большую способность выдержать на длинние периоды в почве вне своих хозяина, через образование структур сопротивления вызванных chlamydospores. Три группы в роду являются наиболее конкретными примерами этих изменений, названных в честь телеморфной группы комплекса Gibberella fujikuroi (Sawada) Wollenw, Fusarium solani (Mart.) Sacc., и комплекса F. graminearum.
Род может представлять колонии цветов от фиолетового, темного пурпуры до оранжевого, но преобладание бледной колонии быстрого роста, воздушного и рассеянного мицелия (MARTINS, 2005). Puhalla (1981); Martins (2005); Agrios (2005), сообщают, что конидии структур (микро и макро), морфология, расположение, разнообразие кониофоров и хламидоспоров были основой для идентификации Fusarium sp. видов. Классические таксономисты описывают наличие одноклеточной и одноклеточной микроконидии и частой многоклеточной макроконидии с клеткой только с одним ядром в качестве генетически идентичного прародителя. Сексуальными стадиями Fusarium являются аскомицеты; сексуальная споры ascospore. Те, выделенные в род Nectria имеют двухклеточные аскоспоры.
Каждая клетка двухклеточного аскопора неуклеизнен, и оба ядра генетически идентичны. Другие, как и в Gibberella, образуют многоклеточные аскоспоры. Martins (2005) упоминает, что морфологические характеристики находятся под влиянием окружающей среды и питания, поэтому культурные средства используются для выбора микроорганизмов и их группировать, исходя из потребностей в питании, которые предоставляются в культурной среде в стандартизированных условиях для их идентификации. Из-за этой пластичности и вариаций в фенотипических характеристиках, найденных в этом грибе, таксономия, основанная только на морфологических понятиях, не является надежной. (MARTINS, 2005; URBENS et al., 2009; QUERALES, 2010).
Идентификация видов Fusarium является одним из первых шагов для его изучения. Традиционно она проводится на основе морфологии, с момента публикации трактата рода “Die Fusarien” в 1935 году Wollenweber; Reinking11. Были предложены и другие системы, но они не решили проблему идентификации видов. Были увеличены инструменты идентификации, включающие использование оптической и электронной микроскопии, селективных и дифференциальных средств, ферментативных сопоставлений, вторичных метаболитов, а также использование иммунологических и молекулярных технологий LEAL-BERTIOLI, 1998; MARTINS 2005). O’Donnell et al., (2000); Summerell et al., (2003); Ottoni, (2008); прокомментировал, что с появлением молекулярных инструментов идентификация Fusarium основана на морфологии, биологии и характеристике вида, основанной на сексуальной совместимости, филогении и последовательностях геномных регионов.
На протяжении десятилетий классическая идентификация использует морфологические характеристики в качестве основы, но вызвала споры, потому что некоторые таксономисты и фитопатологи считают необходимым использовать другие модели для разделения вида, в то время как другие считают, что эти модели влияют на таксономию рода. Примечательно, что практика диагностики причинных агентов заболеваний при посадке на внедрение генетических моделей более адаптирована к исследованиям, проводимым микологами. Тем не менее, эти изменения актуальны, так как в роду могут быть очень близкие виды, и это может привести к ошибочным диагнозам с важными практическими последствиями (QUERALES, 2010).
По словам Notaro et al. (2013), Fusarium solani Math (Sacc) вид, который является преобладающим, вызывает корневую гниль в маниока cv branquinha в штате Pernambuco. Этот вид производит, в среде культуры, воздушный мицелий, с цветом, начиная от белого до крема, становится голубовато-коричневый, когда sporodochios присутствуют. Он производит три типа бесполых спор: слёс-образная макроконидия, с приблизительными размерами 15-35 х 4-7 µm с 3-5 септой; микрокондии, которые имеют или не перегородки, мера около 5-10 х 1-3 µm и chlamydospores, которые гиалин, globose, гладкой грубой, измерения 6-10 µm.
Сексуальное состояние гриба Nectria haematococca (Ascomycete) Этот грибок выживает на инфицированных культурных останков, почвы, клубней и семян, и может сохраняться в течение нескольких лет. Идеальными условиями для вашего учреждения являются температура от 15 до 25°C градусов по Цельсию, влажность от 50 до 75%, возникновение травм, посадка в загрязненных почвах и недостаточное хранение. Повреждения корневой системы, особенно вызванные нематодами, способствуют проникновению гриба в корни хозяев.
3.4 БОРЬБА С БОЛЕЗНЯМИ
Поскольку это возбудитель, переносимый почвой, его ликвидация после установки в посевной зоне становится очень сложной задачей, и химических мер контроля практически не существует, за исключением профилактических, таких как использование семенной обработки или других материалов распространения (LAZAROTTO et al., 2012). Культурный контроль, основанный на профилактике травм во время сбора урожая и хранения, использование здоровых семян и субстрата, свободного от возбудителя, используется только один раз, сбалансированное применение питательных веществ, эффективность в орошении, а также разновидности, которые представляют резистентные характеристики и лечение черенков (манива) и субстрата с фунгицидом для посева при инфицировании , может минимизировать производственные потери.
В качестве контрольной меры наиболее эффективной мерой было основано исследование, проведенное на северо-востоке компанией Embrapa Cassava и Fruit And Agricultural Development Company (EMAGRO), было использование толерантного сорта (Оссо Дюро, Сединья, Бибиана, клон 148/02, Арамарис и Киририс), связанных с растениеводством и системой посадки, что позволило сократить гниль примерно на 80% (GOMES; LEAL, 2003; BETTIOL, 2009; LOBO JÚNIOR et al., 2009; LAZAROTTO et al., 2012).
Для контроля Fusarium sp., химический дифенохоназол и тирам, при обработке почвы и семян, являются наиболее рекомендуемыми из-за эффективности представлены. Однако химический контроль стал постоянным и при неизбирательном использовании возникли экологические проблемы, так как они вызывают изменения в окружающей среде в пользу фитопатогенов с сопротивлением из-за селективного давления, появления вторичных вспышек, сокращения популяции полезных микроорганизмов, из-за отложения резида, который накапливается в почве, воде и пище, делая их вредными, с пагубным воздействием на животных и людей (BIZI, 2015).
В целом высокие затраты на практику химического контроля, связанные с появлением устойчивости к широко используемым продуктам, а также с запрещением бромистого метила, обуновляются разработкой альтернативных методов контроля (BETTIOL, 2009; LOBO JÚNIOR et al., 2009; LAZAROTTO et al., 2012). Для биологической, индукция устойчивости растений является перспективной альтернативой, поскольку она использует натуральные экстракты с микробными и/или индуцирующих свойствами (MORAES, 1992; STANGARLIN et al., 2008).
3.5 БИОКОНТРОЛЬ ФИТОПАТОГЕНОВ
Попытка уменьшить вредные действия патогена или инокулума была постоянной в сельском хозяйстве, и замена агрохимикатов натуральными продуктами с помощью подавления или контрольной деятельности была одной из жизнеспособных альтернатив, поскольку оставшиеся остатки быстрее усваиваются и разлагаются окружающей средой из-за ее низкой стойкости (SOUZA, 2013; BIZI, 2015).
Ethur, (2006) e Dianese, (2007); сообщают о важности исследований в области биологического контроля, таких как введение антагонистических организмов в средах, где патоген наносит ущерб. Cancela (2015), заявляет, что динамика действия биологического контроля происходит естественным образом и состоит в регулировании количества растений и животных естественными врагами. Эта стратегия является основополагающей частью баланса природы и того, как химический контроль имеет преимущества и недостатки. Среди преимуществ является тот факт, что это нетоксичная мера, не вызывает дисбаланса, не имеет противопоказаний, обеспечивает обширный и эффективный контроль, когда нет химического контроля, потому что она будет нуждаться в больше технологии из-за его медленного эффекта, трудно приобрести и не всегда может быть применена в любое время года.
Поддержание баланса в Агроэкосистеме является практикой биологического контроля, чтобы патосистема не нанесла существенного ущерба в биоконтроляционном действии непатогенных организмов системы. Авторы также сообщают, что болезнь больше, чем патоген / принимающих взаимодействия отношения, потому что влияние окружающей среды, а также разнообразие непатогенных организмов лежит в месте инфекции может ограничить или увеличить агрессивность патогена, или сирота сопротивления. Таким образом, патогенные, принимающие и антагонисты взаимодействуют друг с другом в компонентах биологического контроля (BIZI, 2015; CANCELA 2015).
Ограничивающим фактором для использования биологических продуктов является наличие на рынке этого механизма контроля. Другая проблема заключается в том, что многие натуральные продукты не регистрируются должным образом для использования в коммерческих масштабах (SILVA et al., 2008). Присутствие фитопатогена препятствует гомеостазу растений, но оно также может действовать, запуская защитные механизмы и/или активирующие гены, которые придают сопротивление растения-хозяина (AGRIOS, 2005). Michereff et al., (1993),, утверждают, что возбудитель имеет определяющие виды деятельности, связанные с ростом, инфекционностью, агрессивностью, вирулентностью и другими атрибуциями патогена или процессов, которые определяют инфекцию, развитие симптомов и размножение. Взаимодействие между антагонистом, хозяином и патогеном происходит не определенным образом, а взаимно.
Некоторые авторы уже описали способность биоконтрангеров взаимодействовать и вмешиваться в развитие фитопатогена такими механизмами действия, как конкуренция за пространство и питательные вещества к антибиозу, микопаразитизму или гиперпаразитизму, хищничество, индукция сопротивления, тем самым увеличивая конкурентное преимущество в окружающей среде. Эти механизмы варьируются от вида к виду, а также от линии к линии в пределах одного и того же вида, в соответствии с принимающей-паразит взаимодействия. (CARVALHO, 2006; BETTIOL; MORANDI, 2009; BRITO et al., 2010; ZUCCHI, 2010; DIAS, 2011; MACHADO et al., 2012; BIZI, 2015; CANCELA, 2015; FIPKE, et al., 2015).
Многие микроорганизмы обладают высоким биоконтроллером потенциала и рода Trichoderma Pers. был использован в нескольких исследованиях. T. hamatum (Bonord.) Bainier, T. viride Pers., T. aureoviride Rifai, T. harzianum Rifai, T. koningii Oudem, T. pseudokoningii Rifai и ТT. longibrachiatum Rifai являются наиболее цитируемыми (BETTIOL; GHINI, 2005). Ezziyyani et al., (2007) сообщают, что частота Trichoderma spp., объясняется его универсальность либо конкуренции, гиперпаразитизм и антибиоз фитопатогенов, таких как Phytophthora, Fusarium и Neoscytalidium. Remuska & Pria (2007) оценки антагонистического эффекта Trichoderma spp., в контроле микелиального роста фитопатогенных грибов наблюдается, что это оказывает антагонизм на некоторые корень фитопатогенов, таких как Sclerotium rolfsii, Pythium aphanidermatum и Fusarium solani, последний род описывается как один из основных фитопатогенов, связанных с гниливой корня маниоки в штате Пернамбуко.
Hoffmann et al., (2015) сообщили, что они 80% контроль Fusarium sp., используя Trichoderma spp изолирует. Да Silva et al., (2011); Pereira (2013) доказал в своих работах большой уровень ингибирования микелиального роста F. solani видами Trichoderma spp. Souza et al., (2015) также имели многообещающие результаты с Fusarium sp., и Bipolaris sp., используя такой же биоконтроллер. Другие исследования Bomfim et al., (2010); Dias (2011); Lazarotto et al., (2012); Santos et al., (2012); Marques et al., (2014), получил многообещающие результаты, представляющие хорошую отличную производительность, в том что касается ингибирования микелиального роста тестируемых ими фитопатогенов.
3.6 СТРАТЕГИИ БИОКОНТРОЛЯ ТРИХОДЕРМЫ
Род Trichoderma, принадлежащий кСтвивы Королевства, phylum Ascomycota, классы Eauscomycetes и Pyrenomycetes, заказать Hypocreales, семья Hypocreaceae, род Hypocrea (телеоморф). Этот грибок имеет большой антагонистический потенциал и часто ассоциируется с подавляющим почв. (MELO, 1991). По словам Harman et al. (2004); Harman (2005), его использование может предложить и другие преимущества: разложение органических веществ, конкуренция с пагубной микрофлорой через колонизацию ризосферы, производство антибиоза и индукцию резистентности.
Trichoderma видов уже были задокументированы для контроля различных патогенов, таких как Rhizoctonia solani Kühn, Sclerotium rolfsii Sacc., Sclerotina sclerotiorum (Lib.) de Bary, Fusarium spp. e Pythium spp. (MELO; AZEVEDO, 1998). Многочисленные виды Trichoderma, в том числе T. harzianum Hifai, показали высокий уровень тунеядства и антагонистической активности для фитопатогенов, таких как S. rolfsii, S. minor и S. sclerotiorum, R. solani и Pyricularia grisea Sacc. (ROLLAN et al., 1999; RONDÓN et al., 2007).
Согласно Dos Santos (2010), изолят T. harzianum представил антагонизм in vitro над грибком S. rolfsii; возможность гиперпаразитизировать гифы патогена и конкурировать за пространство и питательные вещества. Silva et al. (2013), работая с изолятами Trichodema spp., как биологический агент контроля S. lignicola, отметил, что первоначально изолировать Neoscytalidium sp. это было очень многообещающим, когда сталкиваются с Trichoderma spp., но когда они были найдены в чашке Петри, Trichoderma spp., накладывается патогена колонии. Наблюдая за областью, где антагонист перекрывается с фитопатогенами, можно было заметить деградацию гифы патогена ферментативным действием, так как он точно отличался только спорами и гифами от антагониста.
Для Robbs (1992) и Barbosa; Meza (2009) конкуренция за пищу и антибиоз, являются механизмами наиболее часто используемых биоконтроля агентов, так как размножение и быстрая колонизация являются атрибутами организмов, которые используют имеющиеся ресурсы более эффективно и быстро, обеспечивая их выживание. По словам Ethur(2006), конкурс является одной из основных стратегий Trichoderma sp. изолятов, для этого они используются в качестве биоконтроллеров.
Секреция органических антибиотиков (антибиоза), вырабатываемых микроорганизмами, которые даже в низкой концентрации вмешиваются в метаболическую деятельность, ингибирование или прекращение роста и спорообразуляции, уменьшение прорастания спор и искажения в гифахе и эндолизе, будь то специфические или широкоотравственные, являются важными устройствами для эффективности антагонистов организма (LOBO JUNIOR; ABREU, 2000; BOMFIM, 2007; SOUZA, 2013). Согласно Claydon et al., (1987), антибиотики являются продуктами вторичного метаболизма их производителей, и могут быть более важными в ингибировании других организмов, чем конкуренция за питательные вещества. По сообщениям ряда исследователей, антибиоз также является одним из механизмов действия вида Trichoderma (BETTIOL, 1991; DIAS, 2011; CARVALHO FILHO, 2013). Глиотоксин, виридин и трикодермин являются одними из веществ, производимых видами Trichoderma , способными препятствовать развитию других грибов (Amorim et al., 2011). Ethur (2006) сообщил, что глиотоксин был ответственен за ингибирование прорастания споманджио и микелиальный рост уль Pythium ultimum.
Согласно Bomfim et al., (2010) и Souza (2013), биоконтроллер будет успешен если он мог ухудшить стенку клетки (составленную углеводов, как chitin), также, как протеины и липиды в значительно количестве phytopathogen или прерывают процессы отнесенные к своему синтезу. Таким образом, производство ферментов, которые гидролизуют компоненты стенок клеток фитопатогенов, особенно хитиназы и глюканасы, является общей чертой во многих биоконтроля агентов. Некоторые представители зятя Trichoderma упоминаются в работах как отличный производитель целлюлазы и хитиназы, ферментов, которые деградируют целлюлозу и хитин (MELO, 1991; HARMAN et al., 2004).
Микопараситизм, ситуация, в которой антагонист микроорганизмов живет на или в антагонистизированных грибов, кормления и развития за его счет, является еще одним важным механизмом действия в качестве атрибута хорошего биоконтроллера. Микопаразитизм включает в себя антибиоз и каннибализм, вызванные действием гидролитических ферментов, таких как хитины, глюканасы, протеазы и липазы, которые приводят к смерти одного из них, который, таким образом, служит пищей для выживших (LOBO JUNIOR; ABREU, 2000; HARMAN, 2000; BOMFIM, 2007; BARRA et al., 2008; ZUCCHI, 2010).
Читины и β-1,3-глюканазы непосредственно участвуют во взаимодействии микопаразитизма между видами Trichoderma и их хозяевами, из-за большинства фитопатогенных грибов, обладающих в их стенке клеток хитина, организованных в регулярно упорядоченных слоях и β-1,3-глюканах, расположенных в виде заполнения аморфным образом. Chitinase действует случайным образом и деградирует биологические структуры и компоненты грибковых клеток, выпуская олигомеры, которые вызывают экзокитиназы, тем самым начиная микопаразитическую атаку, гидролизуя олигосахариды стенки клеток фитопатогена. (GIESE et al., 2003; MARCELLO, 2008; BAUERMEISTER et al., 2010).
Melo (1996) сообщает, что некротрофический микопараситизм эффективен в антагонизме к фитопатогенам, особенно с резистентными структурами, которые считаются трудно атакованными микроорганизмами, такими как спросы, склеротии, хламидоспоры и микросклеротии. Melo; Azevedo (1998) утверждает, что Trichoderma sp. он имеет характеристики микопаразита, потому что они могут найти и обнаружить фитопатогены, возможно, биохимические стимулы, расти к гифы восприимчивых грибов, и когда они вступают в контакт, они образуют структуры, такие как аппрессорий, свернуться, в полной мере, проникают и переваривают их. Род Trichoderma, несомненно, является отличным микробным агентом контроля, потому что он представляет, в дополнение к навыкам, описанным выше, основные характеристики, в том, что: воздействие на окружающую среду и низкий или нулевой, легкое распространение (SPIEGEL; CHET, 1998), способность выживать в неблагоприятных условиях, помимо содержащих популяции патогенных микроорганизмов, живущих в почве (VINALE et al., 2008).
4. ЗАКЛЮЧИТЕЛЬНЫЕ СООБРАЖЕНИЯ
Межвидовые экологические отношения – это события, происходящие на различных уровнях в природе и в основном состоящие из деммонических форм, в состоянии взаимосвязи между микроорганизмами, являясь конкуренцией, антибиозом и тунеядством наиболее частыми. Таким образом, болезнетворные микроорганизмы в растениях не только наносят экономические потери, но и развиваются одновременно, нанося тем самым ущерб от начальной стадии развития сельскохозяйственных культур. Были использованы традиционные методы контроля над фитопатогеном, применение химических пестицидов и фунгицидов, также является очень частой формой, которая вызывает серьезные проблемы из-за воздействия на окружающую среду.
Один из наиболее перспективных способов добиться сокращения использования этих агрохимикатов, или даже будущей замены, биологического контроля с помощью многочисленных биоконтроллеров, таких как грибы рода Trichoderma, которые являются эффективными и не вызывают разрушительного воздействия на окружающую среду.
ССЫЛКИ
AGRIOS, G.N. Plant pathology. Amsterdam : Elsevier Academic Press. 5.ed, 2005. 922p.
ALVES, A., A., C.. Fisiologia da mandioca. In: Aspectos socioeconômicos e agronômicos da mandioca. EMBRAPA Mandioca e Fruticultura Tropical, Cruz das Almas, 2006., 817p.
AMORIM, L.; REZENDE, J. A. M.; BERGAMIN FILHO, A. Manual de Fitopatologia. Piracicaba: Agronômica Ceres. v. 1,4. Ed, p. 383 -387, 2011.
BARBOSA, R.J.F.; MEZA, C.L.S. Antagonismo in vitro de Trichoderma harzianum rifai sobre Fusarium oxysporum schlecht f. sp passiflorae en maracuyá (Passiflora edulis sims var. flavicarpa) del municipio zona bananera colombiana. Rev. Fac. Nal. Agr. Medellín, v.62, n.1, p.4743-4748, 2009.
BARRA, V. R.; ROMEIRO, R.S.; FERRAZ, H.G.M.; MACAGNAN, D.; SILVA, H. S. A.; MOURA, A. B.; HALFELD-VIEIRA, B. A; MENDONÇA, H. L.; VIEIRA JÚNIOR, J. R. Potencialidade antagonística em alguns procariotas agentes de biocontrole de enfermidades de plantas. Ver. Summa Phytopathologica, v.34, n.2, p.121-126, 2008.
BAUERMEISTER, A.; REZENDE, M. I.; GIESE, E. C.; DEKKER, R. F. H.; BARBOSA, A. M. 1,3-Glucanases Fúngicas: produção e aplicações biotecnológicas. LONDRINA SP. rev. Semina: Ciências Exatas e Tecnológicas, , v. 31, n. 2, p. 75-86, jul./dez. 2010. http://www.uel.br/seer/index.php/semexatas/article/view/5249
BETTIOL, W. Controle biológico de doencas do filoplano. In: BETTIOL, W., org. Controle biológico de doencas de plantas. Jaguariuna: EMBRAPA-CNPDA, 1991. p.33-52 (EMBRAPA-CNPDA. Documentos, 15).
BETTIOL, W.: MORANDI, M.A.B. Controle Biológico de Doenças de Plantas no Brasil. In: BETTIOL, W.: MORANDI, M.A.B. (Org). Biocontrole de Doenças de Plantas: uso e perspectivas. Jaguariúna – SP: Embrapa Meio Ambiente, p. 7-14, 2009.
BETTIOL, W.; GHINI, R. Solos Supressivos. In: MICHEREFF,S. J.; ANDRADE, D. E. G. T.; MENEZES, M. (Eds.) Ecologia e Manejo de patógenos radiculares em solos tropicais. Recife:UFRPE, Imprensa Universitária, 2005. cap.6, p. 125-143.
BIZI, R. M., Utilização de fungos no Controle Biológico de Doenças. < http://www.floresta.ufpr.br/alias/lpf/public_html/contbio03.html.> Acesso em 16/11/2015.
BOMFIM, M. P., Antagonismo in vitro e in vivo de Trichoderma spp. a Rhizopus stolonifer em maracujazeiro amarelo. Vitória da Conquista BA, 2007. 74f.: Dissertação de Mestrado. Universidade Estadual do Sudoeste da Bahia. 2007
BOMFIM, M. P., SÃO JOSÉ, A. R., REBOUÇAS, T. N. H., ALMEIDA, S. S. D., SOUZA, I. V. B., & DIAS, N. O . Avaliação antagônica in vitro e in vivo de Trichoderma spp. a Rhizopusstolonifer em maracujazeiro amarelo. Summa Phytopathologyca, v.36, n.1, p. 61-67, 2010.
BRITO, F. S.; MILLER, P. R. M., STADNIK, M. Presença de Trichoderma spp em composto e suas características para o controle de fitopatógenos. Revista. Bras. de Agroecologia, Porto Alegre , p. 43-53. 2010.
CANCELA, K. C., Controle Biológico Florestal Conceitos, Terminologia, Descrição e Caracterização de Agente. Proteção florestal Disponível em <http://www.floresta.ufpr.br/alias/lpf/public_html/contbio01.html> Acesso em 16/11/2015.
CARVALHO FILHO, M. R., Relações filogenéticas, identificação e potencial de uso de isolados de Trichoderma no controle do mofo branco e como promotores de crescimento do feijoeiro. Brasília-DF, 2013. 123f. Tese de Doutorado. Programa de Pós-graduação em Fitopatologia, Universidade de Brasília. Brasília. 2013.
CARVALHO, A. C., Bioprospecção de isolados de Trichoderma stromaticum para o controle biológico da Vassoura-de-Bruxa do Cacaueiro. 2006. 82f. Dissertação de Mestrado. Universidade Estadual de Santa Cruz. 2006.
CONAB– CLAYDON, N.; ALLAN, M.; ITANSON, J.R.; AVENT, A. G. Antifungal alkyl pyrones of Trichoderma harzianum. Transactions of the British Mycological Society, v.88, p.503-513, 1987. COMPANHIA NACIONAL DE ABASTECIMENTO. Perspectivas para a agropecuária. Conab, Brasília , v.1, 2013., 154p.
DA SILVA, D. C. V., TIAGO, P. V., & DE SOUZA-MOTTA, C. M. . Isolamento e seleção de fungos filamentosos do solo de sistemas agroflorestais do Município de Bom Jardim (PE) com base na capacidade de produção de enzimas hidrolíticas1. Revista Brasil. Bot, 34(4), 607-610. 2011
DIANESE, A. de C. Variabilidade e controle de Phytophthora palmivora (Podridão-do-pé) e controle da varíola (Aperisporium caricae) do mamoeiro (Carica papaya). 2007. 109f. Tese Doutorado – Universidade de Brasília, Brasília. 2007
DIAS, P. P., Controle biológico de fitopatógenos de solo por meio de isolados de fungos do gênero Trichoderma e sua contribuição no crescimento de plantas. 2011. 101 f. Tese Doutorado. Universidade Federal Rural do Rio de Janeiro. Seropédica – Rio de Janeiro. 2011
DINIZ, M. de S.. Efeito da manipueira na adubação da mandioca. Revista Raízes e Amidos Tropicais, v. 5, 2009.
DOS SANTOS, J. et al. Atividade hiperparasíticade Trichoderma spp. sobre escleródios de Sclerotinia sclerotiorum em solo. In: SEMINÁRIO DE INICIAÇÃO CIENTÍFICA E TECNOLÓGICA,7,2010.Anais…EPAMIG,2010.Online.Disponívelem:http://www.epamig.br/index.phpoption=com_docman&task=cat_view&gid=121&dir=DESC&order=date&limit=10&limitstart=10.Acesso em Dezembro 2015
ETHUR, L.Z. Dinâmica populacional e ação de Trichoderma no controle de fusariose em mudas de tomateiro e pepino. 2006. 154p. Tese (Doutorado em Fitopatologia) – Universidade Federal de Santa Maria, Santa Maria-RS. 2006.
EZZIYYANI, M.; REQUENA, M. E.; EGEA-GILABERT, C.; CANDELA, M. E. Biological Control of Phytophthora Root Rot of Pepper Using Trichoderma harzianum and Streptomyces rochei in Combination. Journal of Phytopathology , [S.I], v.155, n.6, p. 342-349, 2007
FAO. Faostatidtics Database. Disponivel em <https://www.fao.org.br/FAO_Brf2mpu3a.asp> Acesso em Dezembro de 2015.
FIPKE, G. M; PAZINI, J.B.; ETHU,L. Z. Antagonismo de isolados de Trichoderma spp. ao Sclerotinia sclerotiorum em diferentes temperaturas. Magistra, Cruz das Almas BA, V. 27, N.1, p. 23 – 32, Jan./Mar. 2015
FUKUDA C.; OTSUBO A.A. Cultivo da mandioca na região centro sul do Brasil. Embrapa Mandioca e Fruticultura. HTTP://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Mandioca/mandioca_centrosul/doenc s.htm . Acessado em 16 de julho de 2015.
FRAIFE FILHO G. A., BAHIA J. J. S., Mandioca. Ceplac-Cepec-BA. http://www.ceplac.gov.br/radar/mandioca.htm . Acessado Dezembro de 2015.
GIESE, E. C.; BARBOSA, A. M.; SILVA M.L.C. Glucanases Fúngica: Produção e aplicações das -1,3 e -1,6 glucanases. Revista Biotecnologia Ciência e Desenvolvimento, Brasília-DF, nº 30 , P.97-104.. jan/jun 2003.
GOMES, J. C.; LEAL, E. C., Cultivo da Mandioca para a Região dos Tabuleiros Costeiros. In.: Podridão Radicular. Embrapa Mandioca e Fruticultura. Sistemas de Produção, 11. ISSN 1678-8796 Versão eletrônica/Jan/2003. https://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Mandioca/mandioca_tabcosteiros/doencas.htm . Acesso 13/12/2015.
HOFFMANN, C. A. et al.. Potencial de antagonismo de isolados de Trichoderma sp. contra o isolados de Fusarium sp., in vitro. Revista Verde de Agroecologia e Desenvolvimento Sustentável, v. 10, n. 1, p. 236-242, 2015.
HARMAN, G. E. The Nature and Application of Biocontrol Microbes II: Trichoderma spp. Overview of Mechanisms and Uses of Trichoderma spp. Phytopathology, St Paul, v. 96, p. 190-194. 2005
HARMAN, G. E. Myth and dogmas of biocontrol changes in perceptions derived from research on Trichoderma harzianum T-22. Plant Disease, St. Paul, v. 84, p.377-393, 2000.
HARMAN, G.E. HOWELL, C. R., VITERBO, A., CHET, I., & LORITO, M. Trichoderma species-opportunistic, avirulent plant symbionts. Nature Reviews Microbiology. V.2, p. 43-56. 2004.
IBGE – Instituto Brasileiro de Geografia e estatística. PAS – Pesquisa Anual de Serviços, 2015. [online]Disponível na internet via WWW URL: http://www.ibge.gov.br/home/. Arquivo consultado Dezembro de 2015.
LAZAROTTO, M.; BOVOLINI, M. P.; MACIEL, C. G.; MUNIZ, M. F. B. Seleção in vitro de Isolados de Trichoderma spp. com Potencial de Antagonismo a Isolados Patogênicos de Fusarium spp. In: XVI Simpósio De Ensino, Pesquisa E Extensão, Unifra, v. 3, 2012.
LEAL- BERTIOLI, S. C. DE M. O enfoque molecular na sistemática de fungos. Revisão Anual de Patologia de Plantas. V.6. p. 197-230, 1998
LOBO JÚNIOR, M. e ABREU, M.S. Inibição do crescimento micelial de Sclerotinia sclerotiorum por metabólitos voláteis produzidos por alguns antagonistas em diferentes temperaturas e pH’s. Ciência Agrotécnica, Lavras, v.24, p.521-526, 2000.
LOBO JUNIOR, M.; BRANDÃO, R. S.; CORRÊA, C. A.; GÖRGEN, C. A.; CIVARDI, E. A.; OLIVEIRA, P. de. Uso de braquiárias para o manejo de doenças causadas por patógenos habitantes do solo.Santo Antônio de Goiás: Embrapa Arroz e Feijão, 2009. 8 p. (Embrapa Arroz e Feijão. Comunicado técnico, 183).
MACHADO, D. F. M.; PARZIANELLO, F. R.; SILVA, A. C.F. e ANTONIOLLI, Z. I. Trichoderma no Brasil: o fungo e o bioagente. Revista de Ciências Agrárias, v.35, n.1, p. 274-288, 2012.
MARCELLO, C. M. Avaliação da expressão e caracterização de uma exo-b-1,3-glucanase envolvida no mecanismo de micoparasitismo de Trichoderma asperellum. Brasília-DF, 2008. 89f. Tese de Doutorado. Laboratório de Enzimologia Departamento de Biologia Celular do Instituto de Biologia da Universidade de Brasília – UnB. Brasília – DF. 2008.
MARQUES, H. I. P.; SILVA, M. B.; MARQUES, M. D. P.; RODRIGUES, R. C.; RIBEIRO, P. R. C. C. Inibição do crescimento micelial de Trichoderma harzianum por fertilizantes líquidos. Enciclopédia Biosfera, v. 10, n. 18, p. 2040, 2014.
MARTINS, M. K.. Variabilidade genética de isolados de Fusarium spp. e estudo da interação com a planta hospedeira. 2005. 110f. Diss. Tese (Doutorado em Agronomia)-Escola Superior de Agricultura” Luiz de Queiroz”, Universidade de São Paulo, SP.[Links], 2005
MASSOLA JR, N.S.; BEDENDO, I. P.. Doenças da mandioca. In: KIMATI, H. et al. Manual de fitopatologia: Doenças das plantas cultivadas;. 4ª Ed.vol. 2, p. 340-341 – São Paulo: Agronômica Ceres, 2005
MELO, I. S. Potencialidades da utilização de Trichoderma spp. no controle biológico de doenças de plantas. In: BETTIOL, W. (Ed.). Controle biológico de doenças de plantas. Jaguariúna: CNPDA/EMBRAPA, 1991. p. 135-156.
MELO, I. S.; AZEVEDO, J. L. Ecologia Microbiana. Jaguariúna: Embrapa DNPMA, p. 393-419, 1998.
MELO, I.S. Trichoderma e Gliocladium como bioprotetores de plantas. Revis. Anu. Patol. Plantas v. 4, p. 261-295. 1996.
MENEZES, J. P., LUPATINI, M., ANTONIOLLI, Z. I., BLUME, E., JUNGES, E., & MANZONI, C. G. Variabilidade genética na região its do rDNA de isolados de trichoderma spp.(Biocontrolador) e Fusarium oxysporum f. sp. Chrysanthemi. Ciência e Agrotecnologia, 34(1), 132-139.2010
MICHEREFF, S.J.; MENEZES, M.; MARIANO, R.L.R. Potencial de Trichoderma para o controle da antracnose do sorgo. Fitopatologia Brasileira, Brasília, v. 18, p. 392-398, 1993.
MILANESI, P.M. Caracterização, toxicidade e patogenicidade de Fusarium spp. em genótipos de soja em sistema plantio direto. 2009. 91 p. Dissertação (Mestrado em Agronomia) – Universidade Federal de Santa Maria, Santa Maria, 2009.
MORAES, W. B. C. Controle alternativo de fitopatógenos. Pesquisa Agropecuária Brasileira, v. 27, n. 13, p. 175-190, 1992.
MORAIS, M. dos S.; NASCIMENTO, L. C do; MOREIRA, K. A.; SILVA, M. da; CAVALCANTI, N. T. D. O. Levantamento e avaliação da incidência das doenças da mandioca no estado da Paraíba. Summa Phytopathologica, v. 39, n. 3, p. 204. 2013.
MOURA, G. de M.; SILVA, M. D. O. da. Avaliação de resistência de cultivares de mandioca à podridão de raízes. Embrapa-CPAF/AC. Comunicado Técnico, 4p. Rio Branco 1997.
MUNIZ J. N, RUFFINO-NETTO A, YAMAMURA M, VILLA TCS, ARCENCIO R. Aspectos epidemiológicos da coinfecção tuberculose e vírus da imunodeficiência humana em Ribeirão Preto (SP), de 1998 a 2003. J. Bras. Pneumol., 32(6):529-34, 2006
NASCIMENTO JÚNIOR, N. A. Efeito da casca de mandioca no controle da podridão radicular causada por Phytophthora sp. em mandioca de mesa (Manihot esculenta Crantz) var. Rosinha em ambiente irrigado. Rio Largo-AL,2015.82f. Tese de Doutorado. (Programa de Pós-Graduação em Proteção de Plantas da Universidade Federal de Alagoas. Alagoas. 2015.
NOTARO, K. A. MEDEIROS, E. V., SILVA, C. A. D., & Barros, J. A. Prospecção de fitopatógenos associados á podridão radicular da mandioca em Pernambuco, Brasil. Biosci. J., Uberlândia, v. 29, n. 5, p. 1832-1839, 2013.
NOTARO, K. A. Prospecção de fitopatógenos e caracterização de solos arenosos envolvidos na supressividade ou conducividade da podridão radicular da mandioca, causada por Neoscytalidium lignicola/ Krystal de Alcantara Notaro. Garanhuns-PE, 2012. 111f. Dissertação (PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO AGRÍCOLA). Universidade Federal Rural de Pernambuco – Unidade Acadêmica de Garanhuns, 2012.
O`DONNELL, K.; NIRENBERG, H.I.; AOKI, T.; CIGELNIK, E. A multigene phylogeny of the Gibberella fujikoroi species complex: detection of additional phylogenetically distinct species.Mycoscience, Tokyo, v. 41, p. 61-78, 2000.
OLIVEIRA, L. G. SILVA, A.C.S; SILVA, R. L. ,LIMA, D. W. S & LIMA, G; COSTA. Atividade proteolítica de fungos de solo fitopatogênicos ao Feijão-Caupi [Vigna unguiculata (L.) Walp.] In: III CONAC Congresso Nacional de Feijão-Caupi. Pernambuco. Resumo expandido. 4p. 22-24 de abril Recife Pernambuco 2013.
OLIVEIRA, N. T; J. M. A., UCHÔA, S. C. P., RODRIGUES, G. S., MELVILLE, C. C., & de ALBUQUERQUE, J. D. A. A.. Caracterização e identificação de clones de mandioca produzidas em Roraima para o consumo in natura. Revista Agro@mbiente On-line, v. 5, n. 3, p. 188-193, 2012.
OTTONI, R.J. Analise da incidência de Fusarium spp. toxigenicos e de níveis de fumonisinas em grãos ardidos de milho hibrido. 2008. 54 p. Dissertação (Mestrado em Microbiologia Agrícola) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2008.
PEREIRA, C O.F. Estudo da patogenicidade e controle biológico de Fusarium sp. Com Trichoderma sp. 2013.74f. Dissertação. Universidade de Caxias do Sul. 2013.
PUHALLA, J.E. Genetic considerations of the genus Fusarium. In: Nelson, P.E.; Toussoun, T.A.; Cook, R.J. (Ed) Fusarium: diseases, biology, and taxonomy. Pennsylvania: Pennsylvania State University, v.27, p.291-305, 1981.
QUERALES, P. J.. Caracterização morfológica e genética de Fusarium spp. isolados de sementes e associados à podridão do colmo de milho (Zea mays L.). Diss. Escola Superior de Agricultura “Luiz de Queiroz, 2010.
REMUSKA, A.C.; DELLA PRIA, M. Efeito de Bacillus truringiensis e Trichoderma sp. no crescimento de fungos fitopatogênicos. Publication UEPG: Ciências Exatas e da Terra, Agrárias e Engenharia, Ponta Grossa, v.13, n.3, p.31-36, 2007.
ROBBS, F. C. Controle Biológico de doenças em plantas. In: NETO, A. M. A.; BARAN, C. L. (ed.). Manual de Controle Biológico. Rio de Janeiro: Lidador. 1992. p.46-51.
ROLLAN, MONACO C, NICO A. Efecto de la tempratura sobre la interaccion invitr o entre especies de Tricihoderma y Sclerotinia sclerotiorum, S. minor y Sclerotium rolfsii. Invest. Agr.: Prod. Prot. Veg. 14:1-16. 1999.
RONDÓN T.R, GUTIÉRREZ GR, ZAYAS A.D.P, PÉREZ LA, CUTIÑO Y.L. Efectividad in vitro de Trichoderma harzianum Rifai para el biocontrol de Rhizoctonia solani Kühn y Pyricularia Grisea sacc. aislados en el cultivo del arroz (Oryza sativa l.). Fitosanidad 11: 29-34.2007.
SALLA, D. A., CABELLO, C. Análise energética de sistemas de produção de etanol de mandioca, cana-de-açúcar e milho. Revista Energia na Agricultura. Botucatu, vol. 25, n.2, p.32-53., 2010.
SANTOS, C. C., OLIVEIRA, F. A. de; SANTOS, M. S. dos; TALAMINI, V.; FERREIRA, J. M. S.; SANTOS, F. J. dos., Influência de Trichoderma spp. sobre o crescimento micelial de Thielaviopsis paradoxa. Scientia Plena 8, 047309 www.scientiaplena.org.br . 2012
SEIFERT, K.A. Enhancing the relevance of fungal taxonomy to plant pathology: phylogenetics, molecular diagnostics, and long-term memory1. Symposium contribution. Canadian Journal of Plant Pathology, Otawa, v. 28, p. S280-S287, 2006.
SILVA, A. N.; AZEVEDO, G. B.; ROCHA-SOBRINHO, G. G.; NOVAES, Q. S.; Efeito de produtos químicos e de Trichoderma spp. no controle de Fusarium solani do maracujazeiro. Interciencia, v. 6, p. 398-403, 2014.
SILVA, C. A. D., DE MEDEIROS, E. V., BEZERRA, C. B., DE MORAIS; SILVA, W., DE BARROS, J. A., & DOS SANTOS, U. J. Interferência da incorporação de matéria orgânica no solo no controle da podridão negra da mandioca, causada por Scytalidium lignicola. Biosci. J., Uberlândia, v. 29, n. 6 , p. 1823-1831, Nov./Dec. 2013.
SILVA, C. A. D., Prospecção em fitopatógenicos e avaliação de fontes de matéria orgânica sobre a supressividade da podridão radicular da mandioca. Garanhuns- PE, 2013. 77 f. Dissertação (Mestrado em Produção Agrícola). Universidade Federal Rural de Pernambuco – Unidade Acadêmica de Garanhuns, 2013.
SILVA, H. S. A.; ANDRADE, E. C. Impacto potencial das mudanças climáticas sobre as doenças da mandioca no Brasil. In: Impactos das mudanças climáticas sobre doenças de importantes culturas no Brasil. Jaguariúna: Embrapa Meio Ambiente, p. 263-272, 2011.
SILVA, J. L.; TEIXEIRA, R. N. V. Esporulação e crescimento micelial de Fusarium solani em diferentes meios de cultura e regimes de luminosidade. Revista Agrombiente On-line,v.6,n.1,p.47-52, 2012. Disponível em: <http://revista.ufrr.br/index.php/agroambiente/article/view/604>
SILVA, K. S.; REBOUÇAS, T. N. H.; BOMFIM, M. P.; SILVA, D. S.; SÃO JOSÉ, A. R.; BENETT, C. G. S. Atividade antagônica in vitro de isolados de Trichoderma spp. ao fungo Phytophthora citrophthora. Londrina. Semina: Ciências Agrárias, v. 29, n. 4, p. 749-754, out./dez. 2008.
SILVA, M.B.; NICOLI, A. COSTA, A.S.V.; BRASILEIRO, B.G.; JAMAL, C.M., SILVA, C. A.; PAULA JÚNIOR, T. J.; TEIXEIRA, H. Ação antimicrobiana de extratos de plantas medicinais sobre espécies fitopatogênicas de fungos do gênero Colletotrichum. Revista Brasileira de Plantas Medicinais , Botucatu, v.10, n.3, p.57-60, 2008.
SOUSA, R.M.S.; SERRA, I.M.R.S.; MELO, T.A. Effect of essential oils as an alternative in the control of Colletotrichum gloesporioides in pepper.Summa Phytopathologica , v.38, n.1, p.42-47, 2012.
SOUZA, J. R., Potencialidade de fungicida e agente biológico no controle de requeima do tomateiro. 2013. 63f. Dissertação. Universidade Estadual do Sudoeste da Bahia, Vitória da Conquista. Bahia – BR. 2013.
SOUZA, K. de A.;VAZ, M. C. A.; ALVES, G. C. S.; PAZ -LIMA, M. L.. Confronto in vitro de isolados de Trichoderma spp. com fungos filamentosos. 2p. IV Congresso Estadual de Iniciação Científica do IF Goiano 21 a 24 de setembro de 2015.
SPIEGEL, Y.; CHET, I. Evolution of Trichoderma spp. as a biocontrol agent against soilborne fungi and plant parasitic nematodes in Israel. Integrated Pest Management Review, v. 03, p. 167-175, 1998.
STANGARLIN, J.R. et al. Controle de doenças de plantas por extratos de origem vegetal. Revisão Anual de Patologia de Plantas, v.16, p.265-304, 2008.
URBEN, A.F., et al., Curso taxonomia de Fusarium. Embrapa recursos genéticos e biotecnologia. Brasilia-DF: Embrapa informação tecnólogica, 2009.
VENTURA, J. A. Taxonomia de Fusarium e seus segregados: I- história, meios e procedimentos de cultivo. Revisão Anual de Patologia de Plantas , Passo Fundo, v. 7, p. 271-298, 1999.
VINALE F, SIVASITHAMPARAM, K., GHISALBERTI, E. L., MARRA, R., WOO, S. L., & LORITO, M.. Trichoderma–plant–pathogen interactions. Soil Biology and Biochemistry, v. 40, p.1-10.2008.
ZUCCHI, F. O. Trichoderma sp. em áreas agrícolas visando o controle de doenças fúngicas de raízes de plantas cultivadas. Campinas SP. JV Biotecnologia. 15p. 2010.
ПРИЛОЖЕНИЕ – СНОСНАЯ ССЫЛКА
1. WOLLENWEBER, H. W.; REINKING, O. A.. Die fusarien: ihre beschreibung, schadwirkung und bekämpfung. Berlin: P. Parey, 1935
[1] Кандидат наук в области защиты растений, магистр магистерской степени по производству растений, степень бакалавра и бакалавра биологических наук.
[2] Кандидат наук в области агрономии (сельское хозяйство). Степень магистра в области фитопатологии. Специализация в Дидактике для высшего образования. Выпускной в агрономии.
[3] Кандидат наук в области защиты растений. Степень магистра в области агрономии (ирригация и дренаж). Выпускной в агрономии.
[4] Степень магистра в области орошаемого садоводства. Выпускной в агрономии.
[5] Кандидат технических наук. Степень магистра в области агрономии (производство растений и защита растений). Улучшение в улучшении биологических наук. Выпускной в агрономии.
[6] Выпускник по агрономии.
Отправлено: май 2020 года.
Утверждено: июнь 2020 года.