Mário Cézar de Oliveira [1]
Loyane Bertagnolli Coutinho [2]
INTRODUÇÃO
T. gondii teve seu primeiro registro feito por Charles Nicolle e Louis Manceaux (1908) em células mononucleares do baço e do fígado de roedores africanos (Ctenodactylus gundi) e simultaneamente por Alfonso Splendore (1908) no Brasil identificando o parasito em tecido de coelhos (Oryctolagus cuniculus). Nicolle e Manceaux nomearam o gênero Toxoplasma por apresentar uma forma de arco (grego: toxo, arco; plasma, vida) (KIM e WEISS, 2008). Embora T. gondii apresente uma distribuição mundial, existe apenas uma espécie Toxoplasma gondii descrita no gênero Toxoplasma (DUBEY, 2008).
T. gondii causa infecções congênitas e abortos, bem como infecções oportunistas em indivíduos imunocomprometidos (UNNO; KITOH e TAKASHIMA, 2010). Estima-se que aproximadamente 25-30% da população humana mundial esteja infectada por T. gondii (MONTOYA e LIESENFELD, 2004). Contudo, as prevalências variam muito entre países e, muitas vezes, dentro de um determinado país ou entre diferentes comunidades de uma mesma região (PAPPAS et al., 2009). A baixa soroprevalência (10-30%) foi encontrada na América do Norte, sudoeste da Ásia, norte da Europa e nos países em torno do deserto do Saara na África. Prevalências moderadas (30-50%) foram encontradas em países localizados no centro e no sul da Europa e altas prevalências foram encontradas na América Latina e nos países Africanos tropicais (ROBERT-GANGNEUX e DARDÉ, 2012; FLEGR et al., 2014). No Brasil, 50-80% da população adulta é soropositiva para infecção por T. gondii e a transmissão pela água tem sido implicada como uma das maiores causas de infecção nos humanos (BAHIA-OLIVEIRA et al., 2003; DE MOURA et al., 2006).
MORFOLOGIA
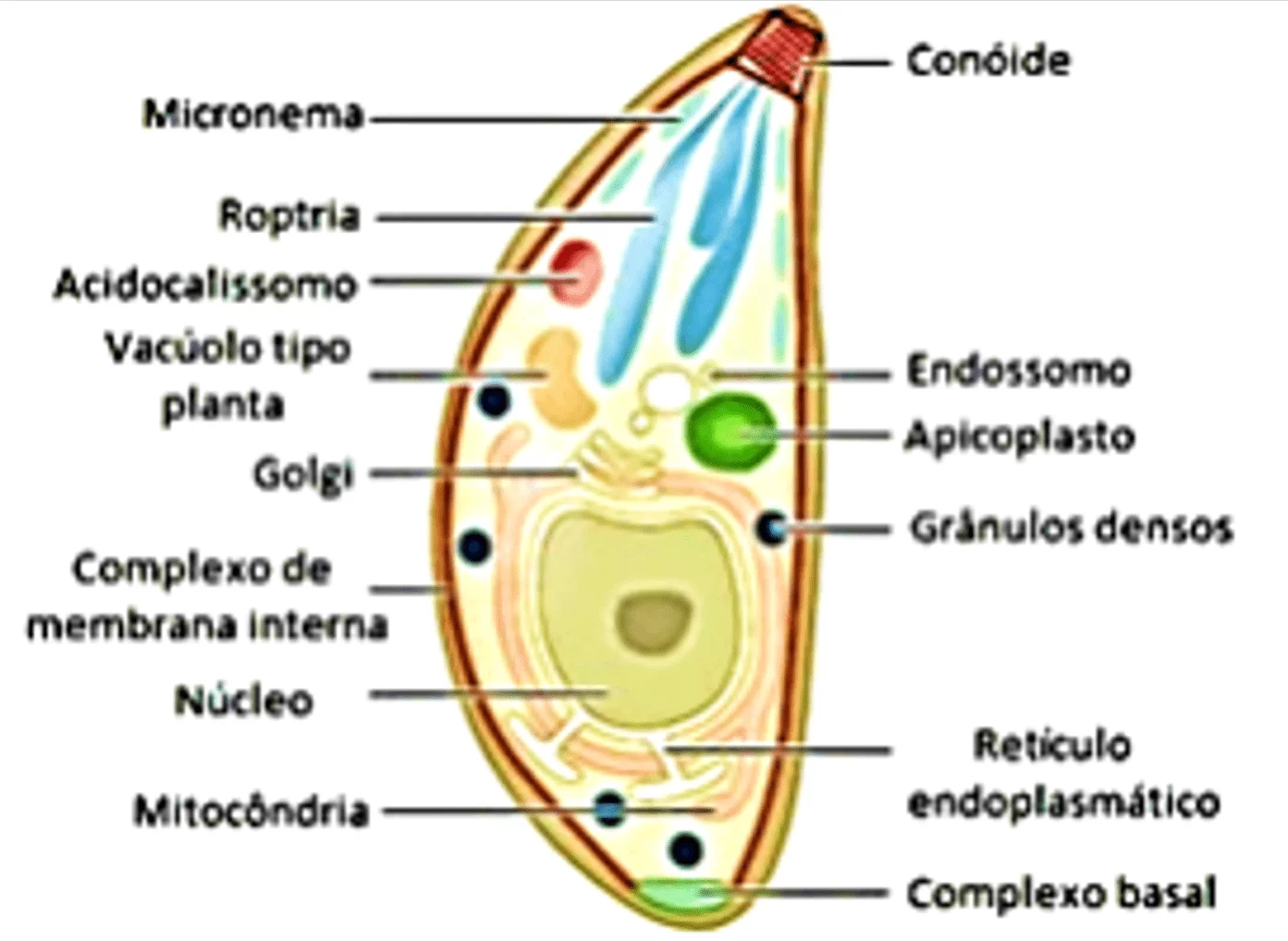
T. gondii possui uma estrutura alongada com forma curvilínea ou crescente com as extremidades arqueadas, podendo também apresentar um formato oval, indicando o início do processo de divisão celular (REY, 2001). Em seu interior encontramos núcleo, situado na região central do parasito ou mais próximo da extremidade posterior, organelas característica de células eucariontes (mitocôndrias, complexo de Golgi, retículo endoplasmático e lisossomo), além de outras organelas particulares ao parasito (conóide, micronemas, roptrias e grânulos densos) (DUBEY et al., 1998; NEVES, 2005) (Figura 1).
A região anterior do parasito é caracterizada pela presença do complexo apical onde se encontra o conóide (Figura 1). O conóide é uma estrutura em forma de cone, formada por três anéis de microtúbulos, sendo dois na região apical e um situado posteriormente, conhecido como anel polar. Do anel polar parte uma rede de citoesqueleto composta por vinte e dois microtúbulos que se dispõe de maneira espiral sob o complexo de membrana interna e se estendem em sentido a região posterior do parasito. No interior do conóide estão as micronemas e roptrias (LERICHE, DUBREMETZ, 1990; NEVES, 2005). Além de manter a forma do parasito, estas estruturas estão envolvidas com a mobilidade e direcionamento de vesículas secretórias durante o mecanismo de invasão à célula hospedeira (DUBEY et al., 1998; HU et al., 2006).
As micronemas são organelas secretoras de moléculas adesivas, domínios tipo integrinas, fator de crescimento epitelial e lectina, sendo localizada na região apical do parasito (FOURMAUX et al., 1996; MEISSNER et al., 2002; SAOUROS et al., 2005) (Figura 1). Essas organelas atuam no reconhecimento e na adesão inicial do parasito aos receptores de superfícies da célula hospedeira (CARRUTHERS, 2002; NEVES, 2005). Onze proteínas de micronemas já foram descritas (MIC1 a MIC11), sendo que a MIC1, MIC4 e MIC6 possuem propriedades de adesão celular (CARRUTHERS, 2002; SIBLEY, 2004).
As roptrias são organelas especializadas associadas com as micronemas, sendo seu conteúdo liberado durante a invasão do parasito, movimento de junção e formação do vacúolo parasitóforo (BRADLEY et al., 2005; BOOTHROYD e DUBREMETZ, 2008). Roptrias estão localizadas na região anterior ao núcleo do parasito e são constituídas por formações membranosas de extremidades dilatadas (LYCKE et al., 1975; BLACK e BOOTHROYD, 2000) (Figura 1). Já foram descritas vinte e nove proteínas de roptrias (ROP1 a ROP29) e elas estão associadas à internalização do parasito à célula hospedeira (BOOTHROYD e DUBREMETZ, 2008). Essa internalização é feita através da liberação de enzimas proteolíticas, como ROP1 que atuam na digestão de proteínas da membrana celular do hospedeiro (KIM e WEISS, 2004). Além disso, as roptrias liberam seu conteúdo proteico no citoplasma da célula hospedeira e atuam na formação do vacúolo parasitóforo, garantindo a natureza não fusogênica do vacúolo e atuando no recrutamento de mitocôndrias e retículo endoplasmático (SINAI et al., 1997; CARRUTHERS, 2002; SINAI, 2008). Outras proteínas tais como Rab11 e toxofilina têm sido identificadas como proteínas pertencentes às roptrias. Rab11 está envolvida no controle de reciclagem do colesterol, enquanto toxofilina se liga à actina facilitando a invasão do parasito (POUPEL et al., 2000; HÖLTTÄ-VUORI et al., 2002).
Além dessas organelas, há no citoplasma dos parasitos, estruturas envoltas por membranas de conteúdo elétron-denso (Figura 1). Tais estruturas denominam-se grânulos densos e contém uma família de glicoproteínas de diferentes pesos moleculares (GRA1 a GRA10). Uma das funções desses grânulos é a liberação de glicoproteínas no vacúolo parasitóforo, possibilitando ao parasito evadir das respostas imunes da célula hospedeira, bem como a formação de túbulos especializados para aumentar a aquisição de nutrientes pelo parasito (COPPENS et al., 2006; CARRUTHERS e BOOTHROYD, 2007).
Figura 1: Representação da estrutura e organelas citoplasmáticas do Toxoplasma gondii
CEPAS DE T. gondii
As cepas de T. gondii são altamente diversas, contudo, apenas algumas linhagens estão amplamente espalhadas. Diferentes genótipos do parasito mostram grande diversidade em patogenicidade e sensibilidade a drogas. Na Europa, América do Norte e África existem três linhagens clonais dominantes de T. gondii conhecidas como tipo I, tipo II e a tipo III (SIBLEY e BOOTHROYD, 1992; FLEGR et al., 2014). Essas linhagens diferem em sua prevalência, virulência, capacidade migratória dentro do hospedeiro e habilidade para converter em bradizoítos. Expansões dessas linhagens coincidem com a domesticação de gatos, bem como, mudanças na prática agrícola (SU et al., 2003). Múltiplas infecções com diferentes cepas podem resultar na produção de um número grande de formas recombinantes e atípicas, que são altamente prevalentes na América do Sul onde cepas exóticas, tais como COUG, MAS e CAST são encontradas (LINDSTRÖM et al., 2008; SIBLEY et al., 2009; DUBEY et al., 2011).
Cepas do tipo I (RH, GT1, CAST e VEL) são virulentas em camundongos e podem causar a morte desses animais. Em humanos, a cepa tipo I está associada a manifestações clínicas graves da toxoplasmose, incluindo manifestações oculares atípicas e surtos agudos, além de ser registradas em pacientes com doenças congênitas (APPLEFORD e SMITH, 2000; GRIGG et al., 2001; BOOTHROYD e GRIGG, 2002).
As cepas do tipo II (ME-49, WIL, PDS e PLK) e tipo III (VEG, MOO, SOU e CEP) são significativamente menos virulentas em camundongos e a infecção por essas cepas tendem a cronificar. Cepa tipo II são frequentes isolados a partir de casos clínicos de toxoplasmose em seres humanos e em pacientes portadores do vírus da imunodeficiência adquirida (HIV). Cepas do tipo III têm sido registradas a partir de isolados de animais (HOWE e SIBLEY, 1995; AJZENBERG et al., 2002).
FORMAS INFECTANTES E CICLO BIOLÓGICO
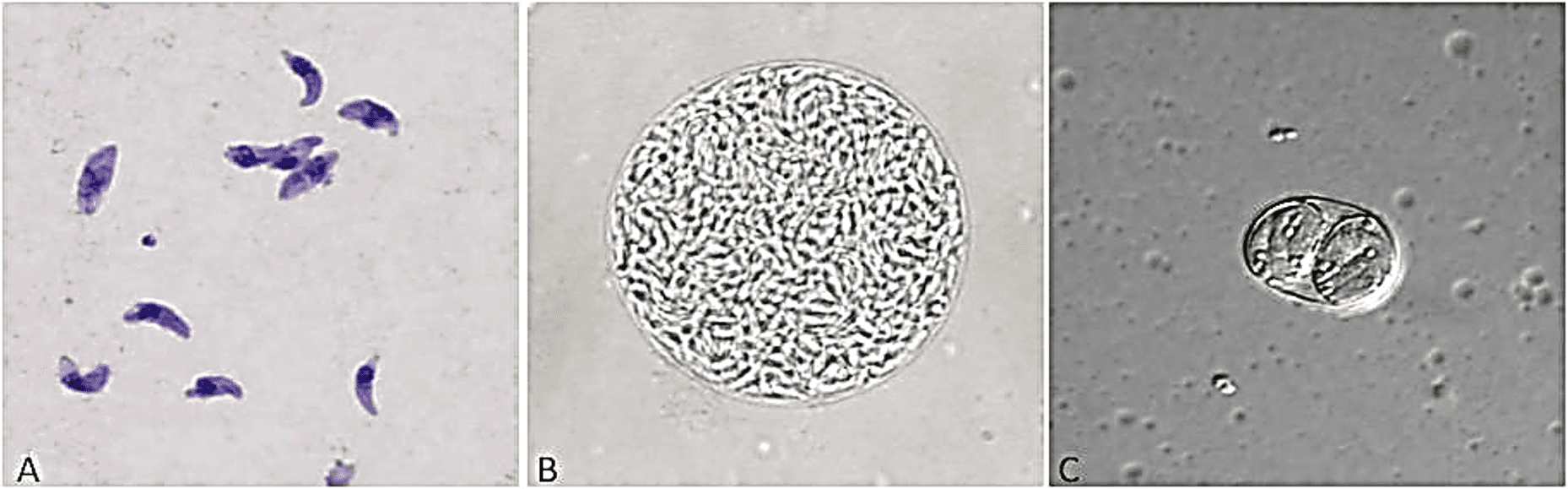
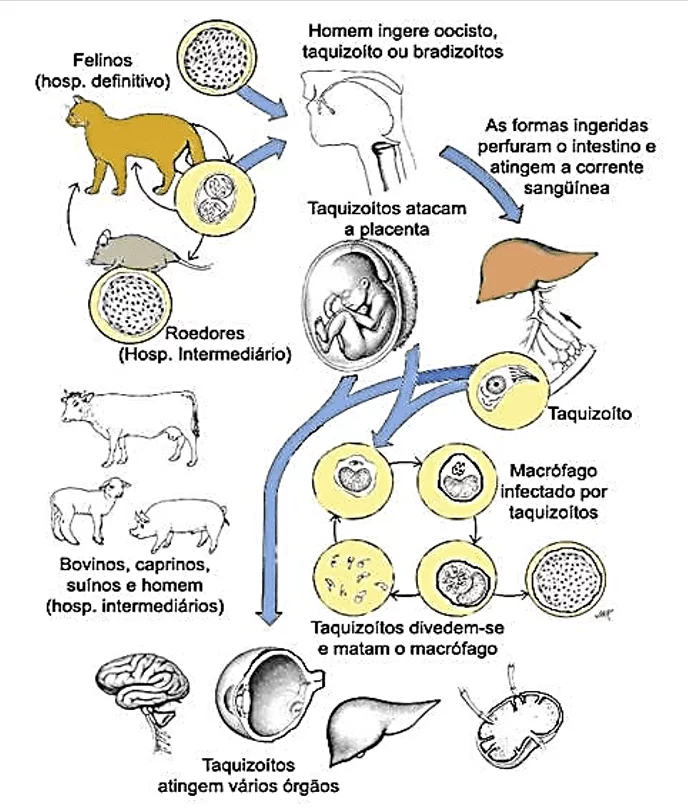
gondii apresenta uma forma de vida heteroxênica, tendo como hospedeiro definitivo (reprodução sexuada) os felídeos e hospedeiros intermediários (reprodução assexuada) animais de sangue quente, incluindo humanos e aves. O parasito apresenta três formas infectantes: taquizoítos, bradizoítos e oocistos (Figura 2) (TENTER et al., 2000; ROBERT-GANGNEUX e DARDÉ, 2012). Todas as três formas são infecciosas tanto para o hospedeiro definitivo quanto para o hospedeiro intermediário, e sua via de transmissão ocorre principalmente por ingestão oral de oocistos infecciosos do meio ambiente (solo, água ou alimentos), ingestão de cistos teciduais em carne crua ou mal cozida e transmissão transplacentária de taquizoítos, além disso, taquizoítos podem ser transmitidos através do leite materno para o filho (DUBEY et al., 1998; CARRUTHERS e SUZUKI, 2007).
O taquizoíto é a forma de disseminação do parasito (grego: tachis = rápido), medindo 2-4 µm de largura e 4-8µm de comprimento, capaz de invadir todo tipo de célula por penetração ativa, multiplicando dentro do vacúolo parasitóforo. Depois de repetidas replicações, as células do hospedeiro são rompidas e os taquizoítos são disseminados pela corrente sanguínea infectando todo o organismo, incluindo o sistema nervoso central, olhos, tecido muscular cardíaco ou esquelético e placenta. Taquizoítos causam uma resposta inflamatória forte, levando a destruição tecidual característica das manifestações clínicas da doença. Sua transformação em bradizoítos ocorre devido à pressão da resposta imune do hospedeiro (DUBEY et al., 1998; MONTOYA e LIESENFELD, 2004) (Figura 2A).
Os bradizoítos são originados a partir da conversão dos taquizoítos em estágios de divisão lenta (grego: brady = lento), são encontrados dentro de cistos teciduais durante a infecção crônica. São morfologicamente idênticos aos taquizoítos, contudo expressam moléculas de estágio específico, apresentando função diferente. Cistos teciduais contêm centenas a milhares de bradizoítos e são formados dentro das células hospedeiras encontradas no tecido muscular cardíaco ou esquelético e cérebro. Seu tamanho é variado podendo medir entre 5 -70 µm. Os bradizoítos podem ser liberados dos cistos e se diferenciarem em taquizoítos causando o reaparecimento da infecção em pacientes imunocomprometidos (DUBEY et al., 1998; MONTOYA e LIESENFELD, 2004) (Figura 2B).
Os oocistos são esféricos, medem entre 10-12µm e possuem uma parede dupla robusta que lhe confere proteção contra ações mecânicas e químicas. Sua parede é formada por mais de 90% de proteínas. Os oocistos são a forma de resistência do parasito e são encontrados nas fezes de felídeos. Durante a infecção aguda, milhares de oocistos são liberados nas fezes dos gatos por 7-21 dias, esses oocistos contém esporozoítos que dão origem aos taquizoítos (DUBEY et al., 1998; MAI et al., 2009; DUMÈTRE et al., 2013) (Figura 2C).
Figura 2: Formas infectantes do Toxoplasma gondii. Taquizoítos (A), Cisto formado por Bradizoítos (B) e Oocisto (C)
A reprodução sexual ocorre somente em felídeos (gatos domésticos ou selvagens). Após a ingestão de cistos presentes no tecido de um hospedeiro intermediário, a parede do cisto é destruída por enzimas gástricas. Os bradizoítos infectam os enterócitos, onde começam a multiplicação assexuada (endodiogenia), caracterizada pelo desenvolvimento de merozoítos dentro do esquizonte (DUBEY, 1998). Esse primeiro passo é seguido por um desenvolvimento sexual com a formação dos gametas masculinos e femininos (gametogonia) no epitélio do intestino delgado (FERGUSON, 2002). Após a fertilização, oocistos formados dentro dos enterócitos, são liberados no lúmen intestinal por rompimento das células e excretados como forma não esporulada nas fezes de gatos (TENTER et al., 2000).
O processo de esporogonia ocorre após alguns dias no meio externo. Mudanças morfológicas levam a formação de um oocisto esporulado com dois esporocistos, cada um contendo quatro esporozoítos haploides. A liberação de oocistos começa 3 a 7 dias após a ingestão de cistos teciduais e pode continuar por mais 20 dias. Gatos infectados podem liberar em torno de 100 milhões de oocistos nas fezes (JONES e DUBEY, 2010). Esses podem infectar vários hospedeiros intermediários, praticamente todos os animais de sangue quente, de mamíferos e aves, quando ingeridos juntos aos alimentos ou água. Oocistos também podem infectar outros gatos embora seja menos eficiente (ROBERT-GANGNEUX e DARDÉ, 2012).
No hospedeiro intermediário, o parasito apresenta duas fases no desenvolvimento assexuado. Na primeira fase, após a ingestão do oocisto, esporozoítos são liberados. Eles penetram no epitélio intestinal onde diferenciam-se em taquizoítos. Esses taquizoítos replicam rapidamente por endodiogenia dentro de qualquer tipo de célula e se disseminam por todo o organismo. A segunda fase começa quando taquizoítos iniciam a conversão para bradizoítos (cisto tecidual) ocorrendo a partir de 7-10 dias após a infecção. Dentro do cisto tecidual, bradizoítos se multiplicam por endodiogenia, e apresentam alta afinidade por tecido muscular e cerebral, contudo, podem ser encontrados em vísceras, tais como pulmão, fígado e rins (DUBEY et al., 1998). Cisto tecidual é o estágio de vida terminal no hospedeiro intermediário sendo altamente infeccioso e pode permanecer durante toda a vida na maioria dos hospedeiros (TENTER et al., 2000).
Após a ingestão desses cistos teciduais por um hospedeiro intermediário através de carne crua ou malcozida, os cistos são rompidos e passam pelo trato digestivo causando a liberação dos bradizoítos. Os bradizoítos irão infectar o epitélio intestinal do novo hospedeiro, podendo se diferenciar em taquizoíto e se disseminar por todo o organismo (ROBERT-GANGNEUX, DARDÉ, 2012) (Figura 3).
Figura 3: Representação esquemática do ciclo biológico do Toxoplasma
INTERAÇÃO PARASITO – HOSPEDEIRO
gondii apresenta a capacidade de invadir uma variedade de células do hospedeiro. Essa invasão é um processo ativo, complexo e depende de múltiplos fatores de regulação, tais como, mobilidade do parasito e secreção de proteínas a partir de suas organelas (micronemas, roptrias e grânulos densos) (BLADER e SAEIJ, 2009; ROBERT-GANGNEUX e DARDÉ, 2012).
A interação com a membrana celular do hospedeiro é um pré-requisito para invasão do parasito e requer a secreção de adesinas dependente de cálcio a partir das micronemas, tais como MIC2, que reconhecem receptores celulares do hospedeiro e promovem a orientação do parasito (WANG et al., 2014). A invasão celular depende de uma interação entre a superfície da célula hospedeira e do parasito, o qual leva a um processo denominado movimentação do tipo gliding, onde ocorre uma interação entre actina-miosina e rearranjos dinâmicos do citoesqueleto do parasito (CARRUTHERS e BOOTHROYD, 2007). Essa invasão é um processo rápido, ocorrendo entre 15 a 30 segundos, sendo distinto do processo normal de endocitose (ROBERT-GANGNEUX e DARDÉ, 2012).
gondii forma uma associação forte entre sua extremidade apical e a membrana da célula hospedeira, o qual ocasiona a internalização do parasito dentro do vacúolo parasitóforo. Esse movimento de junção induz o parasito a expressar em sua superfície o antígeno de membrana apical (AMA1), secretado pelas micronemas, e a secreção de proteínas constituintes das roptrias (RONs), tais como RON2, RON4, RON5 e RON8. Juntas, essas proteínas promovem a locomoção do parasito para o interior da célula hospedeira (ALEXANDER et al., 2005; DUBREMETZ, 2007; STRAUB et al., 2009). A formação da membrana do vacúolo parasitóforo requer a secreção de proteínas das roptrias, tais como ROP-2 e ROP-18, bem como a secreção de proteínas dos grânulos densos durante a primeira hora após a invasão do parasito (EL HAJJ et al., 2007; LALIBERTÉ e CARRUTHERS, 2008).
A maioria das proteínas transmembranas do hospedeiro são removidas da membrana do vacúolo parasitóforo durante o processo de invasão, alterando assim as características bioquímicas da membrana do vacúolo parasitóforo e impedindo a fusão com lisossomos ou qualquer vesícula citoplasmática. A secreção de grânulos densos também mantém o desenvolvimento de uma rede complexa de túbulos de membrana que se desenvolve a partir da membrana do vacúolo parasitóforo e se estende até o lúmen vacuolar (MERCIER et al., 2005).
Essa rede de túbulos está envolvida na troca de nutrientes entre o parasito e a célula hospedeira, facilitando o tráfego de triptofano, arginina, poliamidas, purinas, colesterol, ferro entre outros nutrientes essenciais do citoplasma celular para o parasito, auxiliando em sua sobrevivência (CHAUDHARY et al., 2004; FOX et al., 2004; GAIL et al., 2004; SEABRA et al., 2004; NISHIKAWA et al., 2005; LALIBERTÉ e CARRUTHERS, 2008). Além disso, o vacúolo parasitóforo está associado com as mitocôndrias da célula hospedeira contribuindo para o metabolismo do parasito (ROBERT-GANGNEUX e DARDÉ, 2012). A saída do parasito da célula é um processo ativo que depende do aumento nas concentrações de cálcio intracelular liberado a partir do armazenamento celular (SIBLEY, 2010).
RESPOSTA IMUNOLÓGICA
A resposta imunológica à infecção por T. gondii é individual, complexa e compartimentalizada. Esta variação individual pode ser explicada pelo elevado nível de heterogeneidade no background genético dos indivíduos. Além disso, T. gondii tem a capacidade de se disseminar por todos os tecidos e cada compartimento tecidual apresenta sua própria resposta imune específica, em particular no sistema nervoso central e na placenta (FILISETTI e CANDOLFI, 2004).
Para permanecer no hospedeiro, T. gondii contém uma série de genes que permitem a invasão de células do hospedeiro, modulação da expressão gênica e evasão da resposta imune. Essas estratégias incluem alterações na expressão e secreção de citocinas imunomodulatórias, alterações na viabilidade celular e bloqueio dos mecanismos antimicrobianos. Contudo, T. gondii não consegue suprimir totalmente a resposta imune do hospedeiro, e sim, diminuir parcialmente. Portanto, T. gondii parece manter um delicado equilíbrio entre a indução e supressão da resposta imune, garantindo a sobrevivência tanto do hospedeiro quanto do parasito (MUNOZ et al., 2011).
A defesa do hospedeiro na mucosa intestinal é fornecida pela primeira camada de células epiteliais que revestem o intestino delgado e atuam como uma barreira física impedindo o acesso do conteúdo luminal ao hospedeiro (SHAO et al., 2001). Além disso, as células epiteliais são fontes de quimiocinas que coordenam o recrutamento de células imunes (JU et al., 2009). Células epiteliais especializadas encontradas no intestino delgado, tais como as células de Paneth, são células que liberam defensinas e outros peptídeos antimicrobianos no lúmem intestinal (OUELLETTE et al., 1992; ZASLOFF, 2002). Outro tipo celular encontrado são as células caliciformes, que são células especializadas em secretar glicoproteínas (mucinas), as quais contribuem para a camada de muco protegendo a mucosa epitelial intestinal (ROGERS, 2003).
Subjacente ao epitélio intestinal encontramos a lâmina própria onde residem diversos tipos celulares responsáveis pela defesa contra patógenos, tais como macrófagos, células dendríticas, células Natural Killer (NK), linfócitos T CD4+ e CD8+ e linfócitos B especializados na produção de anticorpo IgA (MENNECHET et al., 2002; VAROL et al., 2010; MASAHATA et al., 2014; WANG et al., 2017). Além disso, células linfóides inatas (ILC) foram identificadas na lâmina própria. Atualmente são conhecidas três populações (ILC1, ILC2 e ILC3) que são definidas pela produção de citocinas imunorreguladoras, que incluem interferon-gama (IFN-y) e interleucinas (IL-4, 5, 13, 17, 22) (SANOS e DIEFENBACH, 2013; KLOSE et al., 2014).
Esses elementos do sistema imune da mucosa intestinal compreende a primeira linha de defesa contra a infecção por T. gondii. Com uma resposta adequada envolvendo uma rede de imunoregulação, a replicação do parasito é controlada e a imunopatologia evitada (COHEN e DENKERS, 2014). As citocinas do perfil T helper 1 (Th1) desencadeiam efeitos microbicidas que matam os taquizoítos, contudo alguns parasitos conseguem evitar a destruição migrando do intestino e estabelecendo infecções latentes sem danos ao hospedeiro (SUZUKI et al., 1988; GAZZINELLI et al., 1991). No entanto, sob inóculos mais elevados ou com certas deficiências imunorreguladoras, T. gondii desencadeia uma resposta imune disfuncional, resultando em inflamação grave, o qual se assemelha à patologia da doença intestinal inflamatória humana (IBD) (EGAN et al., 2012).
Durante a resposta imune inata, monócitos, neutrófilos e células dendríticas são recrutados para os sítios de infecção sendo necessária para resistência do organismo durante a infecção por T. gondii. Uma das funções da resposta imune inata é a capacidade de reconhecer o patógeno e produzir IL-12 que estimulam as células NK e linfócitos T a produzirem IFN-γ (GAZZINELLI et al., 1993; HUNTER et al., 1994). IFN-γ é o maior mediador de resistência à infecção por T. gondii e ativa vários mecanismos intracelulares para eliminar o parasito e inibir sua replicação. Estudos utilizando camundongos geneticamente deficientes em IL-12 ou IFN-γ infectados com T. gondii morrem com a doença aguda e não conseguem controlar a carga parasitária (SUZUKI et al., 1988; GAZZINELLI et al., 1994).
Monócitos estão envolvidos na resistência durante a toxoplasmose, pois essa população celular contribui para o controle direto de T. gondii através da produção de espécies reativas de oxigênio (ROS) e espécies intermediárias de nitrogênio (iNOS), inibindo assim a replicação do parasito (GAZZINELLI et al., 1992; DUNAY e SIBLEY, 2010). Camundongos geneticamente deficientes em iNOS (iNOS-/-) sobrevivem a infecção aguda por T. gondii, contudo, esses animais apresentam uma alta carga parasitária (KHAN et al., 1997).
Durante a resposta imune adaptativa, as células T CD4+ fornecem várias funções reguladoras em mediar à resistência na toxoplasmose. No estágio inicial da infecção, linfócitos T CD4+ contribuem para resposta de linfócitos T CD8+ e linfócitos B através da produção de citocinas, tais como IFN-y ou expressando CD40L, um ligante de CD40 que é expresso em macrófagos e em outras células inatas, ativando mecanismos efetores e fazendo com que essas células controlem a infecção crônica (REICHMANN et al., 2000; JOHNSON e SAYLES, 2002; LÜTJEN et al., 2006). Linfócitos T CD8+ podem controlar a infecção por T. gondii através da produção de citocinas inflamatórias, tais como IFN-y, interação entre CD40/CD40L ou através da citólise mediada pela perforina em células infectadas do hospedeiro (DENKERS et al., 1997; REICHMANN et al., 2000).
Infecção por T. gondii promove a produção de anticorpos e estes podem controlar o parasito. O papel dos anticorpos na imunidade contra T. gondii é demonstrado utilizando camundongos µMT (deficientes em linfócitos B). Esses camundongos desenvolvem uma resposta normal para IFN-y, contudo morrem de 3 a 4 semanas pós-infecção associada com uma alta carga de parasitos no sistema nervoso central (KANG et al., 2000). Esse aumento na suscetibilidade é devido à falta de anticorpos anti-T. gondii, pois um estudo mostrou que a transferência passiva de anticorpos confere proteção em camundongos deficientes em linfócitos B (JOHNSON e SAYLES, 2002). Anticorpos podem mediar efeitos protetores contra T. gondii por vários mecanismos. Estudos in vitro mostraram que esses anticorpos podem opsonizar o parasito para fagocitose, bloquear sua invasão e ativar a via clássica do complemento (SCHREIBER e FELDMAN, 1980; ERBE et al., 1991; VERCAMMEN et al., 1999).
Citocinas reguladoras desempenham um importante papel na resposta contra infecção por T. gondii. A citocina anti-inflamatória IL-10 é altamente regulada durante a infecção oral por T. gondii. A importância desta citocina em prevenir a patologia é evidente pela suscetibilidade de camundongos deficientes em IL-10 causando danos hepáticos e intestinais (GAZZINELLI et al., 1996; SUZUKI et al., 2000). Além disso, camundongos deficientes de IL-10 especificamente em linfócitos T são suscetíveis à patologia intestinal induzida por T. gondii (ROERS et al., 2004).
MANIFESTAÇÃO CLÍNICA
As manifestações clínicas da toxoplasmose são bastante variadas, desde miocardite, toxoplasmose ocular (retinocoroidite, coroidite), encefalite e até mesmo doenças mentais (hidrocefalia) (MORDUE et al., 1999). Porém, as doenças clínicas devido à toxoplasmose não são comuns porque o estágio de taquizoíto (multiplicação rápida), que é responsável pela destruição tecidual e consequente manifestação clínica, é bem controlado pelo sistema imune do hospedeiro. Já a infecção por bradizoítos é latente e, assim, não gera nenhuma doença clínica aparente (BLADER et al., 2015).
No entanto, o parasito atrai considerável atenção médica e científica por provocar doença grave em indivíduos imunocomprometidos, principalmente em pacientes portadores da Síndrome da Imunodeficiência Adquirida (AIDS). Nesses pacientes, a reativação dos parasitos leva ao desenvolvimento da toxoplasmose e a replicação de T. gondii acontece de forma descontrolada, o que provoca danos cerebrais, caracterizados por abcessos e áreas de necrose, com risco de morte. Ao contrário, em indivíduos saudáveis, o parasito é controlado pelo sistema imune e permanece em estado latente podendo, em casos raros, causar uveíte (YAROVINSKY, 2014).
Além dos indivíduos imunocomprometidos, nos quais ocorre a conversão dos bradizoítas em taquizoítas, existe outro grupo de risco que se destaca, sendo ele as mulheres gestantes, nas quais a infecção por T. gondii durante a gestação pode provocar a infecção congênita do feto que, por sua vez, não apresenta um sistema imune maduro, facilitando a replicação descontrolada dos taquizoítos (BLADER et al., 2015) podendo inclusive levar o feto a óbito.
TOXOPLASMOSE CONGÊNITA
Infecções parasitárias congênitas são resultantes da transmissão de parasitos vivos da mulher gestante infectada para o feto os quais persistem após o nascimento. A transmissão pode ocorrer antes do nascimento, no útero (transmissão pré-natal) ou no momento do parto, transmissão perinatal. Tal definição exclui a transmissão de parasitos pós-natal, principalmente a partir do leite materno durante a amamentação, transmissão de parasitos mortos, DNA do parasito ou mesmo moléculas liberadas pelo parasito na mãe e que são prováveis de serem encontradas no sangue fetal. Em uma conotação mais ampla, o termo transmissão vertical corresponde à transmissão materno-fetal de uma geração para outra e inclui transmissão pré-natal, perinatal e, também, rotas de transmissão pós-natal (CARLIER et al., 2012).
As mulheres gestantes representam uma população imunologicamente única uma vez que o sistema imunológico das mesmas é influenciado por sinais originários da placenta, a qual funciona como um órgão imunomodulador, regulando as respostas imunes das células presentes tanto no sítio de implantação quanto sistemicamente. Entretanto, essa modulação não é supressora, mas protetora, pois, geralmente, o sistema imune materno é bem preparado para controlar infecções garantindo, assim, a sobrevivência do feto (PRABHUDAS et al., 2015).
A patogenia da toxoplasmose em humanos é complexa quando a primeira infecção ocorre durante a gestação, sendo que a infecção congênita acarreta, em aproximadamente 30% dos casos, danos cerebrais ou oculares para o feto com aumento da gravidade dependente da idade gestacional, uma vez que as chances de transmissão do parasito aumentam com o avanço da gestação e da prevenção materna (MONTOYA e LIESENFELD, 2004; ROBERT-GANGNEUX et al., 2011).
A infecção dos tecidos placentários pode resultar em placentite e pode levar a uma infecção subsequente das células trofoblásticas, que são um conjunto de células externas da blástula que dará origem ao sinciciotrofoblasto que, por sua vez, irá fazer parte da placenta (ABBASI et al., 2003) sendo que esse processo acarreta duas consequências principais: primeiro que a infecção placentária pode afetar negativamente o equilíbrio tênue da interface materno-fetal e segundo que a placenta pode estar diretamente envolvida na transmissão do parasito para o feto, tornando-se alvo terapêutico e diagnóstico (ROBERT-GANGNEUX et al., 2011).
No microambiente imuneda interface materno-fetal ocorre um balanço Th1/Th2 que exerce influência crucial no desenvolvimento fetal, desempenhando uma importante função na manutenção de uma gestação normal, cujo perfil favorável de resposta imune é o Th2, sendo assim, alterações nesse balanço podem prejudicar a gestação levando ao aborto e fetos natimortos (DEALTRY e O’FARRELL, FERNANDEZ, 2000; ABOU-BACAR et al., 2004).
A citocina IFN-y produzida pelas células NK ou pelos linfócitos citotóxicos CD8+ controla diretamente tanto a invasão dos monócitos e, consequentemente das células trofoblásticas pelo T. gondii, quanto à replicação do parasito nas células infectadas. A liberação exacerbada de IFN-y possui efeitos imunopatológicos tais como apoptose de células teciduais e dilatação da artéria espiral (SENEGAS et al., 2009). Além disso, a secreção de IFN-y provoca alta expressão das moléculas de adesão intracelular (ICAM)-1 na superfície dos trofoblastos aumentando a adesão dos monócitos infectados na superfície das células trofoblásticas, assim, essas células perdem a habilidade de apoptose, o que resulta na persistência do parasito nos tecidos placentários (PFAFF e CANDOLFI, 2008) os quais podem tornar-se um reservatório para a infecção congênita imediata ou tardia.
As células trofoblásticas humanas produzem IL-10 e fator transformador de crescimento beta (TGF-β) (ENTRICAN, 2002) que promovem respostas imunes de perfil Th2 para assegurar a tolerância materno-fetal, por outro lado, tal produção induz aumento significante da invasão e replicação intracelular de T. gondii (BARBOSA et al., 2008). Porém, esses mecanismos imunomoduladores são essenciais para compensar a produção de citocinas Th1 pró-inflamatória induzidas por T. gondii e podem evitar a perda fetal, especialmente quando a infecção ocorre nas fases precoces da gestação (ZHANG et al., 2012a).
O efeito da infecção congênita e o seu grau de acometimento dependerão de vários fatores, como a interação entre o parasito e o hospedeiro, a virulência da cepa do T. gondii, o número de organismos envolvidos (parasitemia), a via de infecção, a capacidade de resposta imune (humoral e celular) do hospedeiro, bem como idade e características genéticas, a integridade das mucosas e das barreiras epiteliais, a idade gestacional na qual a infecção ocorreu e o estágio de maturidade do sistema imune do feto acometido (RORMAN et al., 2006).
Estima-se que, mundialmente, de cada 1000 nascimentos, 3-8 bebês são infectados intrauterinamente e a prevalência de mulheres, em idade gestacional, soropositivas para T. gondii é de 15% nos Estados Unidos da América (EUA), 58% na Europa Central, 51-72% na América Latina e 54-77% em países do Oeste Africano. Baixa soroprevalência, 4-39% foi relatada no Sudoeste da Ásia, China e Coréia, bem como em áreas de clima frio, como países escandinavos (11-28%) e nos EUA 15% das mulheres em idade reprodutiva são soropositivas (TENTER et al., 2000; JONES et al., 2001).
DIAGNÓSTICO
O diagnóstico baseia-se na associação das manifestações clínicas com a confirmação através de estudos sorológicos, ou através da demonstração/detecção do agente etiológico em tecidos ou líquidos corporais em lâminas coradas por Wright-Giemsa ou imuno-histoquímica, a partir de biópsia ou necropsia, testes biomoleculares, ou pela identificação em ensaios experimentais em animais ou em cultivos celulares (LIU et al., 2015). O diagnóstico laboratorial da toxoplasmose tem base na pesquisa de anticorpos contra o parasito. Diferentes marcadores sorológicos têm sido descritos para distinguir entre infecção latente (comum na população) ou infecção recente, como datar na gestante seu contágio pelo T. gondii, ou no imunodeficiente, detectar a reagudização de uma toxoplasmose latente. Devido a essas questões, a sorologia da toxoplasmose se apresenta como das mais complexas, em contínua evolução, exigindo uma variedade de testes que têm sido desenvolvidos com antígenos recombinantes e anticorpos monoclonais dirigidos a regiões cada vez mais restritas e específicas do parasito (FERREIRA e MORAES, 2013; LIU et al., 2015; VAZ; TAKEI e BUENO, 2012).
Uma variedade de testes sorológicos, tais como ELISA (enzyme-linked immunosorbent assay), ensaio de imunoabsorção-aglutinação (ISAGA), teste de anticorpo fluorescente indireto (IFAT) e ensaio de hemaglutinação indireto (IHA) são desenvolvidos para detectar diferentes classes de anticorpos ou antígenos (FERREIRA e MORAES, 2013; LIU et al., 2015; VAZ; TAKEI e BUENO, 2012).
O aumento dos níveis de anticorpos da classe IgG indica a presença de infecção ativa, sendo extremamente importante ser acompanhada da triagem para anticorpos da classe IgM. Níveis de anticorpos IgG baixos e estáveis, podem representar infecção crônica, passada ou persistente. Um teste negativo praticamente descarta uma condição clínica suspeita, sendo necessária uma nova sorologia para descartar a suspeita com oito a dez dias depois do primeiro teste (FERREIRA e MORAES, 2013; LIU et al., 2015, VAZ; TAKEI e BUENO, 2012).
CONSIDERAÇÕES FINAIS
Devido à distribuição cosmopolita e altos índices de infecção pelo T. gondii e a utilização cada vez maior de drogas imunossupressoras, dois grupos de risco para a infecção por este parasito tem sido alvo de estudos: os indivíduos imunossuprimidos e as mulheres gestantes, onde o parasito pode ser transmitido verticalmente e causar alterações importantes ao feto. Adicionalmente, indivíduos imunocompetentes não desenvolvem a doença, sendo necessário que, na vigência da toxoplasmose, esses pacientes sejam avaliados quanto a possível associação com imunodeficientes.
REFERÊNCIAS
ABBASI, M.; KOWALEWSKA-GROCHOWSKA, K.; BAHAR, M. A.; KILANI, R. T.; WINKLER-LOWEN, B.; GUILBERT, L. J. Infection of placental trophoblasts by Toxoplasma gondii. The Journal of Infectious Diseases, v. 188, n. 4, p. 608-16, 2003.
ABOU-BACAR, A.; PFAFF, A. W.; GEORGES, S.; LETSCHER-BRU, V.; FILISETTI, D.; VILLARD, O.; ANTONI, E.; KLEIN, J. P.; CANDOLFI, E. Role of NK cells and gamma interferon in transplacental passage of Toxoplasma gondii in a mouse model of primary infection. Infection and Immunity, v. 72, n. 3, p. 1397-1401, 2004.
AJZENBERG, D.; COGNÉ, N.; PARIS, L.; BESSIÈRES, M. H.; THULLIEZ, P.; FILISETTI, D.; PELLOUX, H.; MARTY, P.; DARDÉ, M. L. Genotype of 86 Toxoplasma gondii isolates associated with human congenital toxoplasmosis, and correlation with clinical findings. The Journal of Infections Disease, v. 186, n. 5, p. 684-9, 2002.
ALEXANDER, D. L.; MITAL, J.; WARD, G. E.; BRADLEY, P.; BOOTHROYD, J. C. Identification of the moving junction complex of Toxoplasma gondii: a collaboration between distinct secretory organelles. PLoS Pathogens, v. 1, n. 2, p. e17, 2005.
APPLEFORD, P. J.; SMITH, J. E. Strain and stage specific variation in Toxoplasma gondii antigens. International Journal for Parasitology, v. 30, n. 11, p. 1187-91, 2000.
BAHIA-OLIVEIRA, L. M.; JONES, J. L.; AZEVEDO-SILVA, J.; ALVES, C. C.; ORÉFICE, F.; ADDISS, D. G. Highly endemic, waterborne toxoplasmosis in north Rio de Janeiro state, Brazil. Emerging Infectious Diseases, v. 9, n. 1, p. 55-62, 2003.
BARBOSA, B. F.; SILVA, D. A.; COSTA, I. N.; MINEO, J. R.; FERRO, E. A. BeWo trophoblast cell susceptibility to Toxoplasma gondii is increased by interferon-gamma, interleukin-10 and transforming growth factor-beta1. Clinical and Experimental Immunology, v. 151, n. 3, p. 536-45, 2008.
BLACK, M. W.; BOOTHROYD, J. C. Lytic cycle of Toxoplasma gondii. Microbiology and Molecular Biology Reviews, v. 64, n. 3, p. 607-23, 2000.
BLADER, I. J.; SAEIJ, J. P. Communication between Toxoplasma gondii and its host: impact on parasite growth, development, immune evasion, and virulence. Acta Pathologica, Microbiologica et Immunologica Scandinavica, v. 117, n. 5-6, p. 458-76, 2009.
BLADER, I. J.; COLEMAN, B. I.; CHEN, C. T.; GUBBELS, M. J. Lytic Cycle of Toxoplasma gondii: 15 Years Later. Annual Review of Microbiology, v. 69, p. 463-85, 2015.
BOOTHROYD, J. C.; GRIGG, M. E. Population biology of Toxoplasma gondii and its relevance to human infection: do different strains cause different disease? Current Opinion in Microbiology, v. 5, n. 4, p. 438-42, 2002.
BOOTHROYD, J. C.; DUBREMETZ, J. F. Kiss and spit: the dual roles of Toxoplasma rhoptries. Nature Reviews: Microbiology, v. 6, n. 1, p. 79-88, 2008.
BRADLEY, P. J.; WARD, C.; CHENG, S. J.; ALEXANDER, D. L.; COLLER, S.; COOMBS, G. H.; DUNN, J. D.; FERGUSON, D. J.; SANDERSON, S. J.; WASTLING, J. M.; BOOTHROYD, J. C. Proteomic analysis of rhoptry organelles reveals many novel constituents for host-parasite interactions in Toxoplasma gondii. The Journal of Biological Chemistry, v. 280, n. 40, p. 34245-58, 2005.
CARLIER, Y.; TRUYENS, C.; DELORON, P.; PEYRON, F. Congenital parasitic infections: A review. Acta Tropica, v. 121, n. 2, p. 55-70, 2012.
CARRUTHERS, V. B. Host cell invasion by the opportunistic pathogen Toxoplasma gondii. Acta Tropica, v. 81, n. 2, p. 111-22, 2002.
CARRUTHERS, V.; BOOTHROYD, J. C. Pulling together: an integrated model of Toxoplasma cell invasion. Current Opinion in Microbiology, v. 10, n. 1, p. 83-9, 2007.
CARRUTHERS, V. B.; SUZUKI, Y. Effects of Toxoplasma gondii infection on the brain. Schizophrenia Bulletin, v. 33, n. 3, p. 745-51, 2007.
CHAUDHARY, K.; DARLING, J. A.; FOHL, L. M.; SULLIVAN, W. J. JR.; DONALD, R. G.; PFEFFERKORN, E. R.; ULLMAN, B.; ROOS, D. S. Purine salvage pathways in the apicomplexan parasite Toxoplasma gondii. The Journal of Biological Chemistry, v. 279, n. 30, p. 31221-27, 2004.
CICLO BIOLÓGICO DO Toxoplasma gondii. Gatos, gravidez e bebês, mito e verdades [Internet]. Acessado em 07/03/2021. Disponível em: http://gatoecachorro.com/gatos-gravidez-e-bebes-mitos-e-verdades/toxoplasmose/.
COHEN, S. B.; DENKERS, E. Y. Border maneuvers: deployment of mucosal immune defenses against Toxoplasma gondii. Mucosal Immunology, v. 7, n. 4, p. 744-52, 2014.
COPPENS, I.; DUNN, J. D.; ROMANO, J. D.; PYPAERT, M.; ZHANG, H.; BOOTHROYD, J. C.; JOINER, K. A. Toxoplasma gondii sequesters lysosomes from mammalian hosts in the vacuolar space. Cell, v. 125, n. 2, p. 261-74, 2006.
DE MOURA, L.; BAHIA-OLIVEIRA, L. M.; WADA, M. Y.; JONES, J. L.; TUBOI, S. H.; CARMO, E. H.; RAMALHO, W. M.; CAMARGO, N. J.; TREVISAN, R.; GRAÇA, R. M.; DA SILVA, A. J.; MOURA, I.; DUBEY, J. P.; GARRETT, D. O. Waterborne toxoplasmosis, Brazil, from field to gene. Emerging Infectious Diseases, v. 12, n. 2, p. 326-9, 2006.
DEALTRY, G. B.; O’FARRELL, M. K.; FERNANDEZ, N. The Th2 cytokine environment of the placenta. International Archives of Allergy and Immunology, v. 123, n. 2, p. 107-19, 2000.
DENKERS, E. Y.; YAP, G.; SCHARTON-KERSTEN, T.; CHAREST, H.; BUTCHER, B. A.; CASPAR, P.; HEINY, S.; SHER, A. Perforin-mediated cytolysis plays a limited role in host resistance to Toxoplasma gondii. Journal of Immunology, v. 159, n. 4, p. 1903-8, 1997.
DUBEY, J. P. Advances in the life cycle of Toxoplasma gondii. International Journal for Parasitology, v. 28, n. 7, p. 1019-24, 1998.
DUBEY, J. P. The history of Toxoplasma gondii-the first 100 years. The Journal of Eukaryotic Microbiology, v. 55, n. 6, p. 467-75, 2008.
DUBEY, J. P.; VELMURUGAN, G. V.; RAJENDRAN, C.; YABSLEY, M. J.; THOMAS, N. J.; BECKMEN, K. B.; SINNETT, D.; RUID, D.; HART, J.; FAIR, P. A.; MCFEE, W. E.; SHEARN-BOCHSLER, V.; KWOK, O. C.; FERREIRA, L. R.; CHOUDHARY, S.; FARIA, E. B.; ZHOU, H.; FELIX, T. A.; SU, C. Genetic characterisation of Toxoplasma gondii in wildlife from North America revealed widespread and high prevalence of the fourth clonal type. International Journal for Parasitology, v. 41, n. 11, p. 1139-47, 2011.
DUBREMETZ, J. F. Rhoptries are major players in Toxoplasma gondii invasion and host cell interaction. Cellular Microbiology, v. 9, n. 4, p. 841-8, 2007.
DUMÈTRE, A.; DUBEY, J. P.; FERGUSON, D. J.; BONGRAND, P.; AZAS, N.; PUECH, P. H. Mechanics of the Toxoplasma gondii oocyst wall. Proceedings of the National Academy of Sciences of the United States of American, v. 110, n. 28, p. 11535-40, 2013.
DUNAY, I. R.; SIBLEY, L. D. Monocytes mediate mucosal immunity to Toxoplasma gondii. Current Opinion in Immunology, v. 22, n. 4, p. 461-6, 2010.
EGAN, C. E.; COHEN, S. B.; DENKERS, E. Y. Insights into inflammatory bowel disease using Toxoplasma gondii as an infectious trigger. Immunology and Cell Biology, v. 90, n. 7, p. 668-75, 2012.
EL HAJJ, H.; LEBRUN, M.; AROLD, S. T.; VIAL, H.; LABESSE, G.; DUBREMETZ, J. F. ROP18 is a rhoptry kinase controlling the intracellular proliferation of Toxoplasma gondii. PLoS Pathogens, v. 3, n. 2, p. e14, 2007.
ENTRICAN, G. Immune regulation during pregnancy and host–pathogen interactions in infectious abortion. Journal of Comparative Pathology, v. 126, n. 2-3, p. 79-94, 2002.
ERBE, D. V.; PFEFFERKORN, E. R.; FANGER, M W. Functions of the various IgG Fc receptors in mediating killing of Toxoplasma gondii. Journal of Immunology, v. 146, n. 9, p. 3145-51, 1991.
FERGUSON, D. J. Toxoplasma gondii and sex: essential or optional extra? Trends in Parasitology, v. 18, n. 8, p. 355-9, 2002.
FERREIRA, A. W.; MORAES, S. L. Diagnóstico laboratorial das principais doenças infecciosas e autoimunes: correlações clínico-laboratoriais. 3ª ed. Rio de Janeiro: Guanabara Koogan, 2013.
FILISETTI, D.; CANDOLFI, E. Immune response to Toxoplasma gondii. Annali dell’Istituto Superiore di Sanità, v. 40, n. 1, p. 71-80, 2004.
FLEGR, J.; PRANDOTA, J.; SOVIČKOVÁ, M.; ISRAILI, Z. H. Toxoplasmosis – a global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS One, v. 9, n. 3, p. e90203, 2014.
FOURMAUX, M. N.; ACHBAROU, A.; MERCEREAU-PUIJALON, O.; BIDERRE, C.; BRICHE, I.; LOYENS, A.; ODBERG-FERRAGUT, C.; CAMUS, D.; DUBREMETZ, J. F. The MIC1 microneme protein of Toxoplasma gondii contains a duplicated receptor-like domain and binds to host cell surface. Molecular and Biochemical Parasitology, v 83, n. 2, p. 201-10, 1996.
FOX, B. A.; GIGLEY, J. P.; BZIK, D. J. Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation. International Journal for Parasitology, v. 34, n. 3, p. 323-31, 2004.
FERREIRA, A. W.; MORAES, S. L. Diagnóstico laboratorial das principais doenças infecciosas e autoimunes: correlações clínico-laboratoriais. 3ª ed. Rio de Janeiro: Guanabara Koogan, 2013.
GAIL, M.; GROSS, U.; BOHNE, W. Transferrin receptor induction in Toxoplasma gondii-infected HFF is associated with increased iron-responsive protein 1 activity and is mediated by secreted factors. Parasitology Research, v. 94, n. 3, p. 233-9, 2004.
GAZZINELLI, R. T.; HAKIM, F. T.; HIENY, S.; SHEARER, G. M.; SHER, A. Synergistic role of CD4+ and CD8+ T lymphocytes in IFN-gamma production and protective immunity induced by an attenuated Toxoplasma gondii vaccine. Journal of Immunology, v. 146, n. 1, p. 286-92, 1991.
GAZZINELLI, R. T.; OSWALD, I. P.; JAMES, S. L.; SHER, A. IL-10 inhibits parasite killing and nitrogen oxide production by IFN-gamma-activated macrophages. Journal of Immunology, v. 148, n. 6, p. 1792-6, 1992.
GAZZINELLI, R. T.; ELTOUM, I.; WYNN, T. A.; SHER, A. Acute cerebral toxoplasmosis is induced by in vivo neutralization of TNF-alpha and correlates with the down-regulated expression of inducible nitric oxide synthase and other markers of macrophage activation. Journal of Immunology, v. 151, n. 7, p. 3672-81, 1993.
GAZZINELLI, R. T.; WYSOCKA, M.; HAYASHI, S.; DENKERS, E. Y.; HIENY, S.; CASPAR, P.; TRINCHIERI, G.; SHER, A. Parasite-induced IL-12 stimulates early IFN-gamma synthesis and resistance during acute infection with Toxoplasma gondii. Journal of Immunology, v. 153, n. 6, p. 2533-43, 1994.
GAZZINELLI, R. T.; WYSOCKA, M.; HIENY, S.; SCHARTON-KERSTEN, T.; CHEEVER, A.; KÜHN, R.; MÜLLER, W.; TRINCHIERI, G.; SHER, A. In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4+ T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha. Journal of Immunology, v. 157, n. 2, p. 798-805, 1996.
GRIGG, M. E.; GANATRA, J.; BOOTHROYD, J. C.; MARGOLIS, T. P. Unusual abundance of atypical strains associated with human ocular toxoplasmosis. The Journal of Infectious Disease, v. 184, n. 5, p. 633-9, 2001.
HÖLTTÄ-VUORI, M.; TANHUANPÄÄ, K.; MÖBIUS, W.; SOMERHARJU, P.; IKONEN, E. Modulation of cellular cholesterol transport and homeostasis by Rab11. Molecular Biology of Cell, v. 13, n. 9, p. 3107-22, 2002.
HOWE, D. K.; SIBLEY, L. D. Toxoplasma gondii comprises three clonal lineages: correlation of parasite genotype with human disease. The Journal of Infections Disease, v. 172, n. 6, p. 1561-6, 1995.
HU, K.; JOHNSON, J.; FLORENS, L.; FRAUNHOLZ, M.; SURAVAJJALA, S.; DILULLO, C.; YATES, J.; ROOS, D. S.; MURRAY, J. M. Cytoskeletal components of an invasion machine-the apical complex of Toxoplasma gondii. PLoS Pathogens, v. 2, n. 2, p. e13, 2006.
HUNTER, C. A.; SUBAUSTE, C. S.; VAN CLEAVE, V. H.; REMINGTON, J. S. Production of gamma interferon by natural killer cells from Toxoplasma gondii-infected SCID mice: regulation by interleukin-10, interleukin-12, and tumor necrosis factor alpha. Infection and Immunity, v. 62, n. 7, p. 2818-24, 1994.
JOHNSON, L. L.; SAYLES, P. C. Deficient humoral responses underlie susceptibility to Toxoplasma gondii in CD4-deficient mice. Infection and Immunity, v. 70, n. 1, p. 185-91, 2002.
JONES, J. L.; KRUSZON-MORAN, D.; WILSON, M.; MCQUILLAN, G.; NAVIN, T.; MCAULEY, J. B. Toxoplasma gondii infection in the United States: seroprevalence and risk factors. American Journal of Epidemiology, v. 154, n. 4, p. 357-65, 2001.
JONES, J. L.; DUBEY, J. P. Waterborne toxoplasmosis-recent developments. Experimental Parasitology, v. 124, n. 1, p. 10-25, 2010.
JU, C. H.; CHOCKALINGAM, A.; LEIFER, C. A. Early response of mucosal epithelial cells during Toxoplasma gondii infection. Journal of Immunology, v. 183, n. 11, p. 7420-7, 2009.
KANG, H.; REMINGTON, J. S.; SUZUKI, Y. Decreased resistance of B cell-deficient mice to infection with Toxoplasma gondii despite unimpaired expression of IFN-gamma, TNF-alpha, and inducible nitric oxide synthase. Journal of Immunology, v. 164, n. 5, p. 2629-34, 2000.
KHAN, I. A.; SCHWARTZMAN, J. D.; MATSUURA, T.; KASPER, L. H. A dichotomous role for nitric oxide during acute Toxoplasma gondii infection in mice. Proceedings of the National Academy of Sciences USA, v. 94, n. 25, p. 13955-60, 1997
KIM, K.; WEISS, L. M. Toxoplasma gondii: the model apicomplexan. International Journal of Parasitology, v. 34, n. 3, p. 423-32, 2004.
KIM, K.; WEISS, L. M. Toxoplasma: the next 100 years. Microbes and Infection, v. 10, n. 9, p. 978-84, 2008.
KLOSE, C. S. N.; FLACH, M.; MÖHLE, L.; ROGELL, L.; HOYLER, T.; EBERT, K.; FABIUNKE, C.; PFEIFER, D.; SEXL, V.; FONSECA-PEREIRA, D.; DOMINGUES, R. G.; VEIGA-FERNANDES, H.; ARNOLD, S. J.; BUSSLINGER, M.; DUNAY, I. R.; TANRIVER, Y.; DIEFENBACH, A. Differentiation of type 1 ILCs from a common progenitor to all helper-like innate lymphoid cell lineages. Cell, v. 157, n. 2, p. 340-56, 2014.
LALIBERTÉ, J.; CARRUTHERS, V. B. Host cell manipulation by the human pathogen Toxoplasma gondii. Cellular and Molecular Life Sciences, v. 65, n. 12, p. 1900-15, 2008.
LERICHE, M. A.; DUBREMETZ, J. F. Exocytosis of Toxoplasma gondii dense granules into the parasitophorous vacuole after host cell invasion. Parasitology Research, v 76, n. 7, p. 559-62, 1990.
LINDSTRÖM, I.; SUNDAR, N.; LINDH, J.; KIRONDE, F.; KABASA, J. D.; KWOK, O. C.; DUBEY, J. P.; SMITH, J. E. Isolation and genotyping of Toxoplasma gondii from Ugandan chickens reveals frequent multiple infections. Parasitology, v. 135, n. 1, p. 39-45, 2008.
LIU, Q.; WANG, Z.; HUANG, S.; ZHU, X. Diagnosis of toxoplasmosis and typing of Toxoplasma gondii. Parasites & Vectors, v. 8, p. 292, 2015.
LÜTJEN, S.; SOLTEK, S.; VIRNA, S.; DECKERT, M.; SCHLÜTER, D. Organ-and disease-stage-specific regulation of Toxoplasma gondii-specific CD8-T-cell responses by CD4 T cells. Infection and Immunity, v. 74, n. 10, p. 5790-801, 2006.
LYCKE, E.; CARLBERG, K.; NORRBY, R. Interactions between Toxoplasma gondii and its host cells: function of the penetration-enhancing factor of Toxoplasma. Infection and Immunity, v. 11, n. 4, p. 853-61, 1975.
MAI, K.; SHARMAN, P. A.; WALKER, R. A.; KATRIB, M.; DE SOUZA, D.; MCCONVILLE, M. J.; WALLACH, M. G.; BELLI, S. I.; FERGUSON, D. J.; SMITH, N. C. Oocyst wall formation and composition in coccidian parasites. Memórias do Instituto Oswaldo Cruz, v. 104, n. 2, p. 281-9, 2009.
MASAHATA, K.; UMEMOTO, E.; KAYAMA, H.; KOTANI, M.; NAKAMURA, S.; KURAKAWA, T.; KIKUTA, J.; GOTOH, K.; MOTOOKA, D.; SATO, S.; HIGUCHI, T.; BABA, Y.; KUROSAKI, T.; KINOSHITA, M.; SHIMADA, Y.; KIMURA, T.; OKUMURA, R.; TAKEDA, A.; TAJIMA, M.; YOSHIE, O.; FUKUZAWA, M.; KIYONO, H.; FAGARASAN, S.; IIDA, T.; ISHII, M.; TAKEDA, K. Generation of colonic IgA-secreting cells in the caecal patch. Nature Communication, v. 5, p. 3704, 2014.
MEISSNER, M.; REISS, M.; VIEBIG, N.; CARRUTHERS, V. B.; TOURSEL, C.; TOMAVO, S.; AJIOKA, J. W.; SOLDATI, D. A family of transmembrane microneme proteins of Toxoplasma gondii contain EGF-like domains and function as escorters. Journal of Cell Science, v. 115, n. 3, p. 563-74, 2002.
MENNECHET, F. J.; KASPER, L. H.; RACHINEL, N.; LI, W.; VANDEWALLE, A.; BUZONI-GATEL, D. Lamina propria CD4+T lymphocytes synergize with murine intestinal epithelial cells to enhance proinflammatory response against an intracellular pathogen. Journal of Immunology, v. 168, n. 6, p. 2988-96, 2002.
MERCIER, C.; ADJOGBLE, K. D.; DÄUBENER, W.; DELAUW, M. F. Dense granules: are they key organelles to help understand the parasitophorous vacuole of all apicomplexa parasites? International Journal for Parasitology, v. 35, n. 8, p. 829-49, 2005.
MORDUE, D. G.; HAKANSSON, S.; NIESMAN, I.; SIBLEY, L. D. Toxoplasma gondii resides in a vacuole that avoids fusion with host cell endocytic and exocytic vesicular trafficking pathways. Experimental Parasitology, v. 92, n. 2, p. 87-99, 1999.
MONTOYA, J. G.; LIESENFELD, O. Toxoplasmosis. Lancet, v. 363, n. 9425, p. 1965-76, 2004.
MUNOZ, M.; LIESENFELD, O.; HEIMESAAT, M. M. Immunology of Toxoplasma gondii. Immunological Reviews, v. 240, n. 1, p. 269-85, 2011.
NEVES, D. P. Parasitologia Humana, 11ª ed. Atheneu, São Paulo, p. 147-56, 2005.
NISHIKAWA, Y.; QUITTNAT, F.; STEDMAN, T. T.; VOELKER, D. R.; CHOI, J. Y.; ZAHN, M.; YANG, M.; PYPAERT, M.; JOINER, K. A.; COPPENS, I. Host cell lipids control cholesteryl ester synthesis and storage in intracellular Toxoplasma. Cellular Microbiology, v. 7, n. 6, p. 849-67, 2005.
OUELLETTE, A. J.; MILLER, S. I.; HENSCHEN, A. H.; SELSTED, M. E. Purification and primary structure of murine cryptdin-1, a Paneth cell defensin. FEBS Letters, v. 304, n. 2-3, p. 146-8, 1992.
PAPPAS, G.; ROUSSOS, N.; FALAGAS, M. E. Toxoplasmosis snapshots: global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. International Journal for Parasitology, v. 39, n. 12, p. 385-94, 2009.
PFAFF, A. W.; CANDOLFI, E. New insights in toxoplasmosis immunology during pregnancy. Perspective for vaccine prevention. Parassitologia, v. 50, n. 1-2, p. 55-8, 2008.
POUPEL, O.; BOLETI, H.; AXISA, S.; COUTURE-TOSI, E.; TARDIEUX, I. Toxofilin, a novel actin-binding protein from Toxoplasma gondii, sequesters actin monomers and caps actin filaments. Molecular Biology of the Cell, v. 11, n. 1, p. 355-68, 2000.
PRABHUDAS, M.; BONNEY, E.; CARON, K.; DEY, S.; ERLEBACHER, A.; FAZLEABAS, A.; FISHER, S.; GOLOS, T.; MATZUK, M.; MCCUNE, J. M.; MOR, G.; SCHULZ, L.; SOARES, M.; SPENCER, T.; STROMINGER, J.; WAY, S. S.; YOSHINAGA, K. Immune mechanisms at the maternal-fetal interface: perspectives and challenges. Nature Immunology, v. 16, n. 4, p. 328-43, 2015.
REICHMANN, G.; WALKER, W.; VILLEGAS, E. N.; CRAIG, L.; CAI, G.; ALEXANDER, J.; HUNTER, C. A. The CD40/CD40 ligand interaction is required for resistance to toxoplasmic encephalitis. Infection and Immunity, v. 68, n. 3, p. 1312-28, 2000.
REY, L. Parasitologia 3ª ed. Rio de Janeiro: Guanabara Koogan S. A., 866p, 2001.
ROBERT-GANGNEUX, F.; DARDÉ, M. L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clinical Microbiology Reviews, v. 25, n. 2, p. 264-96, 2012.
ROERS, A.; SIEWE, L.; STRITTMATTER, E.; DECKERT, M.; SCHLÜTER, D.; STENZEL, W.; GRUBER, A. D.; KRIEG, T.; RAJEWSKY, K.; MÜLLER, W. T cell-specific inactivation of the interleukin 10 gene in mice results in enhanced T cell responses but normal innate responses to lipopolysaccharide or skin irritation. The Journal of Experimental Medicine, v. 200, n. 10, p. 1289-97, 2004.
ROGERS, D. F. The airway goblet cell. The International Journal of Biochemistry & Cell Biology, v. 35, n. 1, p. 1-6, 2003.
RORMAN, E.; ZAMIR, C. S.; RILKIS, I.; BEN-DAVID, H. Congenital toxoplasmosis – prenatal aspects of Toxoplasma gondii infection. Reproductive Toxicology, v. 21, n. 4, p. 458-72, 2006.
SANOS, S. L.; DIEFENBACH, A. Innate lymphoid cells: from border protection to the initiation of inflammatory diseases. Immunology and Cell Biology, v. 91, n. 3, p. 215-24, 2013.
SAOUROS, S.; EDWARDS-JONES, B.; REISS, M.; SAWMYNADEN, K.; COTA, E.; SIMPSON, P.; DOWSE, T. J.; JÄKLE, U.; RAMBOARINA, S.; SHIVARATTAN, T.; MATTHEWS, S.; SOLDATI-FAVRE, D. A novel galectin-like domain from Toxoplasma gondii micronemal protein 1 assists the folding, assembly, and transport of a cell adhesion complex. The Journal of Biological Chemistry, v. 280, n. 46, p. 38583-91, 2005.
SCHREIBER, R. D.; FELDMAN, H. A. Identification of the activator system for antibody to Toxoplasma as the classical complement pathway. The Journal of Infections Disease, v. 141, n. 3, p. 366-9, 1980.
SEABRA, S. H.; DAMATTA, R. A.; DE MELLO, F. G.; DE SOUZA, W. Endogenous polyamine levels in macrophages is sufficient to support growth of Toxoplasma gondii. The Journal of Parasitology, v. 90, n. 3, p. 455-60, 2004.
SENEGAS, A.; VILLARD, O.; NEUVILLE, A.; MARCELLIN, L.; PFAFF, A. W.; STEINMETZ, T.; MOUSLI, M.; KLEIN, J. P.; CANDOLFI, E. Toxoplasma gondii-induced foetal resorption in mice involves interferon-gamma-induced apoptosis and spiral artery dilation at the materno-foetal interface. International Journal for Parasitology, v. 39, n. 4, p. 481-7, 2009.
SHAO, L.; SERRANO, D.; MAYER, L. The role of epithelial cells in immune regulation in the gut. Seminars in Immunology, v. 13, n. 3, p. 163-76, 2001.
SIBLEY, L. D.; BOOTHROYD, J. C. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature, v. 359, n. 6390, p. 82-5, 1992.
SIBLEY, L. D. Intracellular parasite invasion strategies. Science, v. 304, n. 5668, p. 248-53, 2004.
SIBLEY, L. D.; KHAN, A.; AJIOKA, J. W.; ROSENTHAL, B. M. Genetic diversity of Toxoplasma gondii in animals and humans. Biological Science, v. 364, n. 1530, p. 2749-61, 2009.
SIBLEY, L. D. How apicomplexan parasites move in and out of cells. Current Opinion in Biotechnology, v. 21, n. 5, p. 592-8, 2010.
SINAI, A. P.; WEBSTER, P.; JOINER, K. A. Association of host cell endoplasmic reticulum and mitochondria with the Toxoplasma gondii parasitophorous vacuole membrane: a high affinity interaction. Journal of Cell Science, v. 110, n. 17, p. 2117-28, 1997.
SINAI, A. P. Biogenesis of and activities at the Toxoplasma gondii parasitophorous vacuole membrane. Sub-cellular Biochemistry, v. 47, p. 155-64, 2008.
STRAUB, K. W.; CHENG, S. J.; SOHN, C. S.; BRADLEY, P. J. Novel components of the Apicomplexan moving junction reveal conserved and coccidia-restricted elements. Cellular Microbiology, v. 11, n. 4, p. 590-603, 2009.
SU, C.; EVANS, D.; COLE, R. H.; KISSINGER, J. C.; AJIOKA, J. W.; SIBLEY, L. D. Recent expansion of Toxoplasma through enhanced oral transmission. Science, v. 299, n. 5605, p. 414-6, 2003.
SUZUKI, Y.; ORELLANA, M. A.; SCHREIBER, R. D.; REMINGTON, J. S. Interferon-gamma: the major mediator of resistance against Toxoplasma gondii. Science, v. 240, n. 4851, p. 516-8, 1988.
SUZUKI, Y.; SHER, A.; YAP, G.; PARK, D.; NEYER, L. E.; LIESENFELD, O.; FORT, M.; KANG, H.; GUFWOLI, E. IL-10 is required for prevention of necrosis in the small intestine and mortality in both genetically resistant BALB/c and susceptible C57BL/6 mice following peroral infection with Toxoplasma gondii. Journal of Immunology, v. 164, n. 10, p. 5375-82, 2000.
TENTER, A. M.; HECKEROTH, A. R.; WEISS, L. M. Toxoplasma gondii: from animals to humans. International Journal for Parasitology, v. 30, n. 12-13, p. 1217-58, 2000.
UNNO, A.; KITOH, K.; TAKASHIMA, Y. Up-regulation of hyalurona receptors in Toxoplasma gondii-inefected monocytic cells. Biochemical and Biophysical Research Communications, v. 391, n. 1, p. 477-80, 2010.
VAZ, A. J.; TAKEI, K.; BUENO, E. C. Imunoensaios: Fundamentos e Aplicações. [Reimpr.], Rio de Janeiro: Guanabara Koogan, 2012.
VAROL, C.; ZIGMOND, E.; JUNG, S. Securing the immune tightrope: mononuclear phagocytes in the intestinal lamina propria. Nature Reviews. Immunology, v. 10, n. 6, p. 415-26, 2010.
VERCAMMEN, M.; SCORZA, T.; EL BOUHDIDI, A.; VAN BEECK, K.; CARLIER, Y.; DUBREMETZ, J. F.; VERSCHUEREN, H. Opsonization of Toxoplasma gondii tachyzoites with nonspecific immunoglobulins promotes their phagocytosis by macrophages and inhibits their proliferation in nonphagocytic cells in tissue culture. Parasite Immunology, v. 21, n. 11, p. 555-63, 1999.
WANG, Y.; FANG, R.; YUAN, Y.; HU, M.; ZHOU, Y.; ZHAO, J. Identification of host proteins interacting with the integrin-like A domain of Toxoplasma gondii micronemal protein MIC2 by yeast-two-hybrid screening. Parasites & Vectors, v. 7, p. 543, 2014.
WANG, F.; PENG, P. L.; LIN, X.; CHANG, Y.; LIU J.; ZHOU, R.; NIE, J. Y.; DONG, W. G.; ZHAO, Q.; LI, J. Regulatory role of NKG2D+ NK cells in intestinal lamina propria by secreting double-edged Th1 cytokines in ulcerative colitis. Oncotarget, v. 8, n. 58, p. 98945-52, 2017.
WEISS, L. M.; KIM, K. Toxoplasma gondii. The model aplicomplexan: Perspectives and Methods. 2ª ed. Elsevier Science, 800p, 2013.
YAROVINSKY, F. Innate immunity to Toxoplasma gondii infection. Nature Reviews Immunology, v. 14, n. 2, p. 109-21, 2014.
ZASLOFF, M. Antimicrobial peptides of multicellular organisms. Nature, v. 415, n. 6870, p. 389-95, 2002.
ZHANG, R.; ZHANG, H.; LIU, X.; FU, Q.; XU, X.; HU, X. The immunoprotective role of interleukin-10 in abnormal pregnancy outcome induced by Toxoplasma gondii infection. Gynecologic and Obstetric Investigation, v. 73, n. 3, p. 223-9, 2012.
[1] Doutor. ORCID: https://orcid.org/0000-0002-8690-6000. Currículo Lattes: http://lattes.cnpq.br/8924508898024445.
[2] Doutor. ORCID: https://orcid.org/0000-0002-2770-5911. Currículo Lattes: http://lattes.cnpq.br/1698352829926136.



